
| Paleontologia |
|---|
 |
|
Órgãos e processos
|
|
Evolução dos vários táxons
Aves · Borboletas · Cefalópodes · Cetáceos · Dinossauros · Peixes · Fungos · Humanos · Insetos · Mamíferos · Moluscos · Plantas · Répteis · Sirênios · Aranhas · Tetrápodes
|
|
Evolução
|
|
Historia da paleontologia
|
|
Ramos da paleontologia
Bioestratigrafia · Icnologia · Paleontologia dos invertebrados · Micropaleontologia · Palaeoxilologia · Paleobiologia · Paleobotânica · Paleoecologia · Paleogenética · Paleolimnologia · Paleopedologia · Paleotempestologia · Paleozoologia · Palinologia · Esclerocronologia · Tafonomia · Paleontologia dos vertebrados
|
| Portal da Paleontologia |
A evolução dos mamíferos a partir dos sinapsídeos, foi um processo gradual que demorou aproximadamente 70 milhões de anos, do Permiano Médio ao Jurássico Médio. Na metade do Triássico, havia muitas espécies que se pareciam com mamíferos, e apenas no início do Jurássico aparecem os primeiros mamíferos verdadeiros. O primeiro marsupial conhecido, o Sinodelphys, apareceu há 125 milhões de anos, no início do Cretáceo; mais ou menos na mesma época se desenvolvem os Eutheria, e dois milhões de anos mais tarde surgiram os primeiros Monotremos. Na maciça extinção do Cretáceo-Terciário desaparecem os dinossauros, restando as formas aviárias, e os mamíferos iniciam um processo de diversificação e ocupação dos nichos ecológicos vagos, até que no fim do Terciário todas as ordens modernas já se haviam estabelecido.
Do ponto de vista da nomenclatura filogênica, os mamíferos são os únicos sinapsídeos sobreviventes, uma linhagem que se distinguiu dos sauropsídeos, os répteis, no fim do Carbonífero, tornando-se os maiores e mais comuns vertebrados do período Permiano. O desenvolvimento de algumas características típicas dos mamíferos, como a endotermia, os pelos e o cérebro maior, pode ter sido estimulado pelo predomínio anterior dos dinossauros, que ocupavam os nichos ecológicos diurnos e forçaram os mamaliformes para os nichos noturnos.
As evidências dessa evolução se encontram principalmente nos fósseis. Durante um bom tempo fósseis dos mamíferos mesozoicos e seus antecessores imediatos eram escassos e fragmentários, mas o estudo se aprofundou quando na década de 1990 foram encontrados diversos achados importantes, especialmente na China. As novas técnicas científicas como a filogenética molecular também deram contribuição significativa, esclarecendo e fixando pontos de divergência evolutiva que levaram ao surgimento de espécies modernas. Embora as glândulas mamárias sejam a assinatura nos mamíferos modernos, pouco se conhece sobre sua evolução e sobre o processo de lactação, e menos ainda sobre o desenvolvimento do neocórtex, outro traço distintivo desse grupo. O estudo sobre a evolução dos mamíferos se concentra atualmente no desenvolvimento dos ossículos do ouvido médio a partir da articulação da mandíbula dos ancestrais amniotas, junto com a análise da evolução da postura ereta dos membros, do palato secundário, do pelo e do sangue quente.
Definição de "mamífero"
As espécies de mamíferos atuais podem ser identificadas pela presença nas fêmeas de glândulas mamárias que produzem leite.
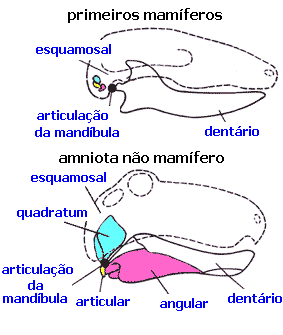
Outras características são necessárias para classificar os fósseis, já que as glândulas mamárias e outros tecidos macios não estão disponíveis.
Os paleontólogos então usam uma característica distintiva que é compartilhada por todos os mamíferos viventes (inclusive os monotremados), mas que não está presente em qualquer dos primeiros terapsídeos triássicos ("mamíferos semelhantes a répteis"): os mamíferos usam dois ossos para ouvir que todos os outros amniotas usam para comer. Os mais antigos amniotas tinham a articulação da mandíbula composta pelo osso articular (um pequeno osso na parte posterior da mandíbula inferior) e pelo osso quadratum (um pequeno osso na parte posterior da mandíbula superior). Todos os não mamíferos amniotas usam este sistema, inclusive os lagartos, os crocodilianos, os dinossauros (e seus descendentes, as aves) e os terapsídeos. Mas os mamíferos têm uma articulação da mandíbula diferente, composta apenas pelo osso da mandíbula inferior que carrega os dentes e pelo esquamosal (outro pequeno osso do crânio). E nos mamíferos, os ossos articular e quadratum se tornaram os ossos bigorna e martelo no ouvido médio.
Os mamíferos também possuem um par de côndilos occipitais - eles têm duas saliências na base do crânio que encaixam na vértebra superior do pescoço, enquanto os outros vertebrados possuem apenas um côndilo occipital. Mas os paleontólogos usam apenas a articulação da mandíbula e o ouvido médio como critérios para identificação de fósseis mamíferos, porque seria confuso se eles encontrassem um fóssil que possuísse uma característica, mas não a outra (por exemplo, uma mandíbula e um ouvido mamíferos, mas um côndilo occipital não-mamífero).
Devido às mudanças incrementadas nos fósseis transicionais, afirma-se
Nós talvez façamos novamente a pergunta: "O que é um mamífero?" Onde nós estipulamos o limite entre réptil e mamífero não tem importância biológica. É puramente uma questão de conveniência. Há duas escolhas óbvias, e ambas imediatamente seguem um período de rápida evolução que torna definitiva a quebra que nós esperamos encontrar.
Os ancestrais dos mamíferos
Aqui, uma "árvore genealógica" bastante simplificada. O texto abaixo descreve algumas das incertezas e áreas de debate.

Amniotas
Os primeiros vertebrados completamente terrestres eram amniotas - seus ovos tinham membranas internas que permitiam ao embrião desenvolvido respirar, mas deixavam que a água entrasse. Isto permitiu aos amniotas pôr seus ovos em terra firme, enquanto os anfíbios geralmente precisavam pôr seus ovos na água (alguns poucos anfíbios, como o sapo do Suriname, desenvolveram outros caminhos para contornar esta limitação).
Os primeiros amniotas aparentemente surgiram no final do Carbonífero a partir de reptiliomorfos ancestrais. Em poucos milhões de anos, duas importantes linhagens amniotas se tornaram distintas: os ancestrais sinapsídeos dos mamíferos e os sauropsídeos, dos quais lagartos, cobras, crocodilianos, dinossauros e aves são descendentes.
Os mais antigos fósseis conhecidos de todos estes grupos datam de cerca de 320 a 315 milhões de anos. Desafortunadamente, é difícil ter certeza sobre quando cada um deles evoluiu, visto que fósseis vertebrados do final do Carbonífero são muito raros, e dessa forma, as primeiras ocorrências reais de cada um destes tipos de animais devem ser consideravelmente anteriores.
Sinapsídeos

Os crânios sinápsidas são identificados pelo padrão distintivo das aberturas temporais atrás de cada olho. Estas aberturas tornam o crânio leve sem sacrificar a força, economizam energia por usar menos osso e provavelmente forneciam pontos de ligação para os músculos da mandíbula. Possuir pontos de ligação fora das mandíbulas tornou possível aos músculos serem mais compridos e então exercer uma forte influência sobre uma grande variedade de movimentos da mandíbula sem serem esticados ou contraídos além de seu melhor alcance.
Os fósseis terrestres do começo do Permiano indicam que um grupo sinápsida, os pelicossauros, foram os vertebrados terrestres mais comuns e incluíam os maiores animais terrestres de seu tempo.
Terapsídeos
Os terapsídeos evoluíram dos pelicossauros na metade do Permiano e tomaram sua posição como vertebrados terrestres dominantes. Eles diferiam dos pelicossauros em várias características do crânio e da mandíbula, incluindo grandes aberturas temporais e incisivos de mesmo tamanho.
Os terapsídeos atravessaram vários estágios, começando como animais que eram muito parecidos com seus ancestrais pelicossauros e terminando como algo que poderia facilmente ser confundido com os mamíferos.
- Desenvolvimento gradual de um osso secundário do palato duro: muitos livros e artigos interpretam isto como um pré-requisito para a evolução da alta taxa metabólica dos mamíferos, porque possibilita a estes animais comer e respirar ao mesmo tempo. Mas alguns cientistas assinalam que alguns animais ectotérmicos modernos usam uma palato secundário feito de carne para separar a boca das vias respiratórias, e que um palato ósseo fornece uma superfície na qual a língua pode manipular a comida, facilitando a mastigação e não a respiração. A interpretação do palato secundário ósseo como uma ajuda para a mastigação também sugere o desenvolvimento de um metabolismo rápido, desde que a mastigação torne possível digerir a comida mais rapidamente. Nos mamíferos, o palato é formado por dois ossos específicos, mas vários terapsídeos permianos tinham outras combinações de ossos nos lugares exatos para funcionarem como palatos.
- O osso dentário gradualmente se tornou o principal osso da mandíbula inferior.
- O progresso na direção de uma postura de membros eretos, o que aumentaria a força animal evitando a restrição de Carrier. Mas esse processo foi instável e muito lento - por exemplo: todos os terapsídeos herbívoros mantiveram membros não eretos (algumas formas posteriores devem ter tido membros traseiros semi-eretos); os terapsídeos carnívoros permianos possuíam membros dianteiros não eretos, e alguns outros do final do Permiano tinham membros traseiros semi-eretos. Na realidade, os monotremados modernos ainda possuem membros semi-eretos.
- No Triássico, avançaram na direção da mandíbula mamífera e do ouvido médio.
- Há evidências plausíveis de pêlos nos terapsídeos do Triássico, mas não nos terapsídeos do Permiano.
- Alguns cientistas argumentam que alguns terapsídeos do Triássico apresentam sinais de lactação.
Árvore genealógica terápsida
Árvore genealógica terápsida; apenas com aqueles que são mais relevantes para a evolução dos mamíferos estão descritos abaixo.

Apenas dicinodontes, terocéfalos e cinodontes eram sobreviventes no Triássico.
Biarmosuchia
Os Biarmosuchia eram os mais primitivos e parecidos com os pelicossauros dos terapsídeos.
Dinocéfalos
Os dinocéfalos ("cabeças terríveis") eram enormes, alguns tão grandes quanto um rinoceronte, e incluíam carnívoros e herbívoros. Alguns dos carnívoros tinham membros posteriores semi-eretos, mas todos os dinocéfalos possuíam membros dianteiros não eretos. Em muitas características, eles eram terapsídeos muito primitivos. Por exemplo, eles não possuíam um palato secundário e suas mandíbulas eram particularmente "reptilianas".
Teriodontes
Os teriodontes e seus descendentes tinham articulações de mandíbulas nas quais o osso articular da mandíbula inferior estava fortemente ligado ao muito pequeno osso quadratum do crânio. Isto permitia uma grande abertura da boca, e um grupo, o dos carnívoros gorgonopsídeos, tirou vantagem disto para desenvolver "dentes de sabre". Mas a mandíbula dos teriodontes teve um significativo longo alcance - o tamanho muito reduzido do osso quadratum foi um passo importante no desenvolvimento da articulação da mandíbula dos mamíferos e do ouvido médio.
Os gorgonopsídeos ainda tinham algumas características primitivas: ausência de palato secundário ósseo (mas outros ossos nos lugares apropriados para executar as mesmas funções); membros dianteiros não eretos; membros traseiros que poderiam funcionar em posturas eretas e não eretas. Mas os terocefalídeos, que aparentemente surgiram aproximadamente ao mesmo tempo dos gorgonopsídeos, tinham características adicionais semelhantes aos mamíferos como, por exemplo, seus ossos dos dedos tinham o mesmo número de falanges dos primeiros mamíferos (e o mesmo número que os primatas têm, inclusive os humanos).
Cinodontes
Os cinodontes, um grupo teriodonte que também surgiu no final do Permiano, inclui os ancestrais de todos os mamíferos - um subgrupo, os tritelodontes, é amplamente considerado em conter provavelmente o ancestral dos mamíferos. As características semelhantes às dos mamíferos dos cinodontes incluem: redução posterior no número de ossos da mandíbula inferior; um palato ósseo secundário; molares com um complexo padrão nas coroas; o cérebro preenchendo a cavidade endocranial.
Tomada de poder triássica: a ascensão dos sauropsídeos
A catastrófica extinção em massa do Permiano-Triássico acabou com cerca de 70 por cento das espécies vertebradas terrestres, assim como a maioria das plantas.
Como resultado:
- Os ecossistemas e as cadeias alimentares desmoronaram, e a recuperação levou cerca de 6 milhões de anos.
- Os sobreviventes recomeçaram a luta pelo domínio de seus antigos nichos ecológicos - até mesmo os cinodontes, que aparentemente estavam no caminho da dominação no final do Permiano.
Mas os cinodontes foram derrotados por um grupo antes obscuro de sauropsídeos, os arcossauros (que incluía os ancestrais dos crocodilianos, dos dinossauros e das aves). Esta mudança de sorte é frequentemente chamada de "tomada de poder triássica". Várias explicações têm sido propostas para ela, mas o mais provável é que o começo do Triássico foi predominantemente árido e então a capacidade de conservação de água superior dos arcossauros deu-lhes uma vantagem decisiva (todos os sauropsídeos conhecidos tinham peles sem glândulas e excretavam ácido úrico, que requeria menos água para mantê-lo suficientemente líquido, que a ureia que os mamíferos excretam e presumivelmente os terapsídeos excretavam). A tomada de poder triássica foi gradual - no começo do Triássico, os cinodontes eram os principais predadores e os listrossauros os principais herbívoros, mas na metade do Triássico, os arcossauros dominaram todos os grandes nichos carnívoros e herbívoros.
Mas a tomada de poder triássica deve ter sido um fator vital na evolução dos cinodontes em mamíferos. Os descendentes dos cinodontes só foram capazes de sobreviver como insectívoros pequenos e principalmente noturnos. Como resultado:
- A tendência terápsida em direção a dentes diferenciados com oclusão precisa se acelerou, por causa da necessidade de segurar os artrópodes capturados e esmagar seus exoesqueletos.
- A vida noturna exigia avanços no isolamento térmico e na termorregulação para possibilitar aos ancestrais dos mamíferos serem ativos no frio da noite.
- Sentidos acurados de audição e olfato se tornaram vitais.
- Isto acelerou o desenvolvimento do ouvido médio mamífero, e então da mandíbula, visto que os ossos que tinham sido parte da articulação da mandíbula se tornaram parte do ouvido médio.
- O aumento no tamanho dos lóbulos olfativo e auditivo do cérebro aumentou o peso cerebral percentualmente em relação ao peso do corpo. O tecido cerebral exige uma quantidade desproporcional de energia. A necessidade de mais comida para manter cérebros maiores aumentou a pressão por melhoras no isolamento térmico, termorregulação e alimentação.
- Como um efeito colateral, a visão se tornou levemente menos importante, e isto se reflete no fato de que muitos mamíferos têm uma visão de cores pobre, incluindo os prosímios como os lêmures.
Dos cinodontes aos verdadeiros mamíferos
Muitas incertezas
Enquanto a tomada de poder triássica provavelmente acelerou a evolução dos mamíferos, ela tornou a vida mais difícil para os paleontólogos, porque bons fósseis do quase-mamíferos são extremamente raros, principalmente porque eles eram majoritariamente menores que ratos:
- Eles estavam amplamente restritos aos ambientes que são menos capazes de fornecer bons fósseis. Os melhores ambientes terrestres para fossilização são as planícies alagadas, onde alagamentos sazonais rapidamente cobrem animais mortos com uma camada protetora de silte que depois é comprimida em uma rocha sedimentar. Mas as planícies alagadas são dominadas por animais médios e grandes, e os terapsídeos triássicos e os quase-mamíferos não poderiam competir com os arcossauros em tamanho.
- Seus ossos delicados eram frágeis para serem destruídos antes que pudessem ser fossilizados - por comedores de animais mortos (incluindo os fungos e as bactérias) e por serem espalhados.
- Pequenos fósseis são difíceis de localizar e mais vulneráveis a serem destruídos pela erosão e por outros fenômenos naturais antes de serem descobertos.
Na realidade, foi dito recentemente, na década de 1980, que todos os fósseis de mamíferos e quase-mamíferos do Mesozoico poderiam estar guardados em algumas caixas de sapatos - e eles eram principalmente dentes, que são os mais duráveis de todos os tecidos.
Como resultado:
- Em muitos casos, é difícil classificar um fóssil mamífero ou quase-mamífero mesozoico em um gênero.
- Todos os fósseis disponíveis de um gênero raramente formam um esqueleto completo, e portanto a dificuldade em se decidir quais gêneros são mais parecidos uns com outros e então mais proximamente relacionados. Em outras palavras, torna-se muito difícil classificá-los através da cladística, que é o método mais confiável e menos subjetivo atualmente disponível.
Dessa forma, a evolução dos mamíferos no Mesozoico é repleta de incertezas, embora não haja espaço para a dúvida de que os mamíferos apareceram pela primeira vez no Mesozoico.
Mamíferos ou mamaliformes?
Um dos resultados destas incertezas foi uma mudança na definição de "mamífero" pelos paleontólogos. Durante muito tempo, um fóssil era considerado um mamífero se nele fosse encontrado o padrão mandíbula-ouvido médio (a articulação da mandíbula consistindo apenas do esquamosal e do dentário; e os ossos articular e quadratum tornando-se os ossos martelo e bigorna do ouvido médio). Mas mais recentemente, os paleontólogos têm geralmente definido "mamífero" como o último ancestral comum de monotremados, marsupiais e placentários e de todos os seus descendentes. E então eles tiveram que definir um outro clado, o dos mamaliformes, para acomodar todos os animais que são mais semelhantes a mamíferos que a cinodontes, mas menos aparentados com monotremados, marsupiais e placentários. Ainda que isto agora pareça ser a voz da maioria, alguns paleontólogos têm resistido, porque: a nova definição simplesmente move a maior parte dos problemas para o novo clado sem esclarecê-los; o clado dos mamaliformes inclui alguns animais com a articulação da mandíbula "mamífera" e outros com a articulação da mandíbula "reptiliana" (articular-quadratum); e a nova definição de "mamífero" e "mamaliformes" é dependente dos últimos ancestrais comuns de ambos os grupos, e que ainda não foram encontrados. Apesar destas objeções, este artigo segue a opinião dominante, e trata a maioria dos descendentes mesozoicos dos cinodontes como mamaliformes.
Árvore genealógica - dos cinodontes aos mamíferos
(Baseada em Mammaliformes - Palaeos)

Multituberculados
Os multituberculados (assim chamados pelos múltiplos nódulos ou saliências - tubérculos - em seus "molares") são muitas vezes chamados de "roedores do Mesozoico", mas trata-se de um exemplo de convergência evolutiva e não significa que eles sejam aparentados com os roedores. À primeira vista eles se assemelham a mamíferos: suas articulações da mandíbula consistem apenas dos ossos dentário e esquamosal e os ossos articular e quadratum fazem parte do ouvido médio; seus dentes são diferenciados e têm cúspides semelhantes às dos mamíferos; eles têm um arco zigomático; a estrutura da pélvis sugere que eles davam à luz a filhotes indefesos muito pequenos, como os modernos marsupiais. Eles viveram por mais de 120 milhões de anos (do Jurássico médio, há cerca de 160 milhões de anos, até o começo do Oligoceno, há cerca de 35 milhões de anos), o que faria deles os mamíferos mais bem sucedidos de sempre. Mas uma olhada mais de perto mostra que eles eram muito diferentes dos mamíferos modernos:

- Seus "molares" têm duas filas paralelas de saliências, diferente dos molares tribofênicos (de três pontas) dos primeiros mamíferos.
- O movimento de mastigação é completamente diferente. Os mamíferos mastigam com um movimento de trituração de lado a lado, o que significa que habitualmente os molares se fecham em apenas um lado ao mesmo tempo. As mandíbulas dos multituberculados eram incapazes do movimento de lado a lado e mastigavam arrastando os dentes inferiores para trás, contra os superiores conforme a mandíbula fechava.
- A parte anterior do arco zigomático geralmente consiste do osso maxilar e não do jugal, que é um osso minúsculo numa pequena fenda na extensão do maxilar.
- O esquamosal não faz parte do crânio.
- O focinho é diferente do dos mamíferos. Na realidade, parece-se mais com o dos pelicossauros como o dimetrodonte. O focinho multituberculado é em forma de caixa, com a enorme e plana maxila formando os lados, o osso nasal acima e o exagerado pré-maxilar na frente.
Morganucodontidae
Os morganucodontes apareceram pela primeira vez no final do Triássico, há cerca de 205 milhões de anos. Eles são um excelente exemplo de fósseis transicionais, visto que possuem tanto articulações da mandibula dentário-esquamosal quanto articular-quadratum. Eles também foram um dos primeiros descobertos e mais perfeitamente estudados dos mamaliformes, por causa de um extraordinário grande número de fósseis morganucodontes encontrados.
Docodontes

O mais conhecido membro dos docodontes é o Castorocauda lutrasimilis, que viveu no médio Jurássico, há cerca de 164 milhões de anos e foi descoberto em 2004 e descrito em 2006. O Castorocauda ("cauda de castor") não foi um docodonte típico (a maioria era onívora) nem exatamente um mamífero, mas é extremamente importante no estudo da evolução dos mamíferos porque o primeiro esqueleto encontrado estava quase completo (um verdadeiro luxo na paleontologia) e quebra o estereótipo do "pequeno insetívoro noturno":
- Ele era perceptivelmente maior em tamanho que a maioria dos fósseis semelhantes a mamíferos do Mesozoico - cerca de 43 centímetros do nariz à ponta de sua cauda de 13 centímetros, e provavelmente pesaria mais que 800 gramas.
- Ele fornece a mais antiga evidência sem nenhuma dúvida de cabelos ou pêlos. Anteriormente, o mais antigo era o Eomaia scansoria, um mamífero verdadeiro de há cerca de 125 milhões de anos.
- Ele tinha adaptações aquáticas que incluíam ossos achatados e restos de tecido macio entre os dedos dos pés traseiros sugerem que eles eram membranosos. Antes disso, os mais antigos animais semi-aquáticos semelhantes a mamíferos eram do Eoceno, há cerca de 110 milhões de anos.
- Os poderosos membros diateiros do Castrocauda parecem adaptados para escavação. Esta característica e as esporas em seus tornozelos fazem-no lembrar o ornitorrinco, que também nada e escava.
- Seus dentes parecem adaptados para comer peixe: os dois primeiros molares têm cúspides em linha reta, o que faz deles mais adequados para agarrar e cortar que para triturar; e estes molares são curvados para trás, para ajudar a capturar presas escorregadias.
Hadrocodium
A árvore genealógica simplificada acima mostra o Hadrocodium como um parente evolutivo mais distante dos mamíferos, enquanto os simetrodontes e os Kuehneotheriidae são mais proximamente relacionados com os verdadeiros mamíferos. Mas os fósseis de simetrodontes e Kuehneotheriidae são tão poucos e fragmentados que eles ainda são pouco conhecidos e talvez sejam parafiléticos. Por outro lado, há bons fósseis de Hadrocodium (de há cerca de 195 milhões de anos, do princípio do Jurássico) e eles possuem algumas características importantes:
- A articulação da mandíbula consiste apenas dos ossos dentário e esquamosal, e as mandíbulas não possuem pequenos ossos na parte de trás do dentário, diferente do modelo terápsida.
- Nos terapsídeos e na maioria dos mamaliformes, o tímpano se estica sobre um canal na parte de trás da mandíbula inferior. Mas o Hadrocodium não tem tal canal, o que sugere que seu ouvido era parte do crânio, como nos mamíferos - e, portanto, que os antigos ossos articular e quadratum migraram para o ouvido médio e se transformaram nos ossos martelo e bigorna. Por outro lado, o dentário tinha uma "entrada" na parte posterior que os mamíferos não têm. Isto sugere que o osso dentário do Hadrocodium manteve a mesma forma que ele teria se os ossos articular e quadratum permanecessem fazendo parte da articulação da mandíbula, e dessa forma, que o Hadrocodium, ou um ancestral muito próximo, deve ter sido o primeiro a ter um ouvido médio completamente mamífero.
- Os terapsídeos e os mamaliformes mais antigos tinham suas articulações da mandíbula em uma posição bastante recuada no crânio, parcialmente porque o ouvido ficava na extremidade posterior da mandíbula, mas também porque tinha que estar próximo do cérebro. Este arranjo limitava o tamanho da caixa craniana, porque forçava os músculos da mandíbula a ficar em torno e sobre ela. A caixa craniana e as mandíbulas do Hadrocodium não eram mais ligados um ao outro pela necessidade de sustentar o ouvido, e sua articulação da mandíbula era mais avançada. Nos seus descendentes ou naqueles animais com um arranjo similar, a cavidade cerebral estava livre para se expandir sem ser limitada pela mandíbula e a mandíbula estava livre para mudar sem ser limitada pela necessidade de manter o ouvido médio próximo do cérebro - em outras palavras, agora se tornava possível para os animais semelhantes a mamíferos desenvolver cérebros grandes e adaptar suas mandíbulas e dentes nas formas que fossem puramente especializadas para comer.
Os mais antigos mamíferos verdadeiros
Esta parte da história introduz novas complicações, visto que os mamíferos verdadeiros são o único grupo que ainda possui membros vivos:
- Uma tem que distinguir grupos extintos daqueles que tem representantes viventes.
- Outra muitas vezes sente-se compelida a explicar a evolução de características que não aparecem nos fósseis. Este esforço às vezes envolve filogenética molecular, uma técnica que se tornou popular a partir da metade da década de 1980, mas é ainda controversa por causa de suas hipóteses, especialmente sobre a confiabiidade do relógio molecular.
Árvore genealógica dos primeiros mamíferos
(baseada em Mammalia: Overview - Palaeos)

Australosphenida e Ausktribosphenidae
Ausktribosphenidae é um nome dado a um grupo de achados particularmente incompreensível que:
- Parece ter molares tribosfênicos, um tipo de dente que é encontrado apenas nos placentários e nos marsupiais.
- São originários de depósitos do Cretáceo médio na Austrália - mas a Austrália estava conectada apenas à Antártida, e os placentários se originaram no hemisfério norte e estiveram confinados a ele até a deriva continental formar pontes de terra entre a América do Norte e a América do Sul, entre a Ásia e a África e entre a Ásia e a Índia (o mapa do final do Cretáceo em [1] mostra como os continentes do sul estavam separados).
- São representados apenas por fragmentos de crânios e mandíbulas, o que não é de muita ajuda.
Australosphenida é um grupo que foi definido para incluir os Ausktribosphenidae e os monotremados. O Asfaltomylos (da metade para o fim do Jurássico), encontrado na Patagônia) é aparentemente um australosfenídeo basal (um animal que: tem características compartilhadas entre Ausktribosphenidae e monotremados; carece de características que são peculiares a Ausktribosphenidae ou monotremados; também não possui características que estão ausentes nos Ausktribosphenidae e monotremados) e mostra que os australosfenídeos eram amplamente disseminados por toda Gondwana (o antigo super-continente do hemisfério sul).
Monotremados
O mais antigo monotremado conhecido é o Teinolophos, que viveu há cerca de 125 milhões de anos na Austrália. Análises recentes sugerem que ele não era um monotremado ancestral, primitivo, mas um ornitorrinco completo, e então que as linhagens do ornitorrinco e do equidna divergiram consideravelmente antes e que os monotremados ancestrais eram anteriores.
Os monotremados têm algumas características que devem ter sido herdadas dos amniotas originais:
- Eles usam o mesmo orifício para urinar, defecar e reproduzir-se ("monotremado" significa "orifício único") - da mesma forma que os lagartos e as aves.
- Eles põem ovos com casca de couro, como aqueles de lagartos, tartarugas e crocodilos.
Diferente dos outros mamíferos, as fêmeas dos monotremados não possuem mamilos e alimentam seus filhotes por leite "suado" de glândulas de seus abdômens.
Naturalmente, estas características não são visíveis nos fósseis, e as principais características do ponto de vista dos paleontólogos são::
- Um osso dentário delgado no qual o processo coronoide é mínimo ou inexistente.
- A abertura externa do ouvido situa-se na base posterior da mandíbula.
- O osso jugal é pequeno ou inexistente.
- Uma cintura escapular primitiva com fortes elementos ventrais: coracoides, clavículas e inter-clavicular (os mamíferos terianos não possuem interclavícula).
- Membros dianteiros não eretos ou semi-eretos.
Theria
Theria ("bestas") é um nome aplicado ao grupo hipotético a partir do qual os grupos metatheria (que inclui os marsupiais) e eutheria (que inclui os placentários) descendem. Embora nenhum fóssil convincente de terianos básicos tenha sido encontrado (apenas uns poucos dentes e fragmentos da mandíbula são conhecidos), os grupos metatheria e eutheria compartilham algumas características que supostamente teriam sido herdadas de um grupo ancestral comum:
- Não possuem inter-clavicular.
- Ossos coracoides não existentes ou fundidos com os omoplatas para formar os processos coracoides.
- Molares tribosfênicos.
- Um tipo de tornozelo crurotarsal onde: a articulação principal está entre a tíbia e o tálus; o calcâneo não tem contato com a tíbia, mas forma um calcanhar ao qual os músculos podem se ligar. (O outro tipo bem conhecido de tornozelo crurotarsal é visto nos crocodilianos e funciona de maneira distinta - a maior parte da curva do tornozelo está entre o calcâneo e o tálus.
Metatheria
O grupo moderno metatheria inclui todos os marsupiais ("animais com bolsas"). Alguns gêneros, tais como o mongol do final do Cretáceo Asiatherium, podem ser marsupiais ou membros de algum outro grupo metatheriano.
O mais antigo marsupial conhecido é o Sinodelphys, encontrado em uma camada de xisto de 125 milhões de anos (começo do Cretáceo) na província de Liaoning, no nordeste da China. O fóssil está quase completo e inclui tufos de pêlos e impressões de tecidos macios.
Os didelfídeos (marsupiais comuns do hemisfério ocidental) apareceram pela primeira vez no final do Cretáceo e ainda têm representantes viventes, provavelmente porque eles são em sua maioria onívoros semi-arborícolas não especializados.
A característica melhor conhecida dos marsupiais é o seu método de reprodução:
- A mãe desenvolve um tipo de saco gema em seu útero que fornece nutrientes ao embrião. Os embriões de bandicoots, coalas e vombates adicionalmente formam órgãos semelhantes a placentas que os conecta à parede uterina, embora estes órgãos sejam menores que nos mamíferos placentários e não há certeza de que eles transfiram nutrientes da mãe para o embrião.
- A gestação é muito curta, tipicamente de quatro a cinco semanas. O embrião nasce precocemente, e é normalmente menor que cinco centímetros no nascimento. Sugere-se que a gestação curta é necessária para reduzir o risco de que o sistema imunológico da mãe ataque o embrião.
- o marsupial recém-nascido usa seus membros dianteiros (com mãos relativamente fortes) para subir até um mamilo, que normalmente está numa bolsa no ventre da mãe. A mãe alimenta o bebê pela contração de músculos sobre suas glândulas mamárias, já que o bebê é muito fraco para sugar. A necessidade do marsupial recém-nascido de usar seus membros dianteiros na subida até o mamilo evitou que estes se desenvolvessem em remos ou asas e, dessa forma, evitou o surgimento de marsupiais aquáticos ou voadores verdadeiros (embora haja vários marsupiais planadores).
Embora alguns marsupiais sejam muito parecidos com alguns placentários (o tilacino é um bom exemplo), os esqueletos masuspiais possuem algumas características que os distinguem dos placentários:
- Alguns, incluindo o tilacino, têm quatro molares. Nenhum placentário tem mais que três.
- Todos têm um par de fenestras palatais, aberturas na parte inferior do crânio (além das pequenas aberturas das narinas).
Os masuspiais também têm um par de ossos marsupiais (chamados às vezes de "ossos epipúbicos"), que sustentam a bolsa nas fêmeas. Mas estes não são exclusivos aos marsupiais, visto que eles foram encontrados em fósseis de multituberculados, monotremados e até mesmo de placentários - de modo que eles são provavelmente uma característica ancestral comum que desapareceu em algum momento depois que o ancestral dos mamíferos placentários modernos divergiu daquele dos marsupiais. Alguns pesquisadores acham que a função original dos ossos epipúbicos era ajudar na locomoção pela sustentação de alguns dos músculos que empurram a coxa para frente.
Eutheria
Os membros do grupo moderno eutheria ("bestas verdadeiras") são todos placentários. Mas o mais antigo eutheriano conhecido, o Eomaia scansoria, encontrado na China e datado de há cerca de 125 milhões de anos, possui algumas características que são mais parecidas com aquelas dos marsupiais (os metaterianos modernos):
- Ossos epipúbicos prolongando-se adiante da pélvis, que não são encontrados em nenhum placentário moderno, mas são encontrados nos marsupiais, monotremados, mamaliformes e multituberculados. Em outras palavras, esses ossos aparentam ser uma característica ancestral que deve ter estado presente nos primeiros placentários.
- Uma abertura pélvica estreita, o que indica que as crias eram muito pequenas ao nascer e dessa forma a gestação era curta, como nos marsupiais modernos. Isto sugere que a placenta foi um desenvolvimento posterior.
- Cinco incisivos em cada lado da mandíbula superior. Este número é típico dos methaterianos, e o número máximo nos placentários modernos é três, com exceção dos homodontes como o tatu. Mas a razão molar-pré-molar do Eomaia (ele possuía mais pré-molares que molares) é típica dos eutherianos (placentários) e não é normal nos marsupiais.
O Eomaia também tinha um canal meckeliano, uma característica primitiva da mandíbula inferior que não é encontrada nos modernos mamíferos placentários.
Estas características intermediárias são consistentes com as estimativas da filogenética molecular de que os placentários se diversificaram há cerca de 110 milhões de anos, 15 milhões de anos depois da datação do Eomaia fóssil.
O Eomaia também tinha muitas características que sugerem fortemente que ele era um escalador, incluindo: várias características dos pés e dedos dos pés; pontos de fixação bem desenvolvidos para os músculos que são bastante usados para escalar; e uma cauda que era duas vezes mais longa que o resto da espinha dorsal.
A característica placentária mais bem conhecida é seu método de reprodução:
- O embrião se prende ao útero por uma grande placenta pela qual a mãe fornece alimento e oxigênio e remove os dejetos.
- A gestação é relativamente longa e os bebês são bastante desenvolvidos ao nascer. Em algumas espécies (especialmente herbívoros que vivem nas planícies), o recém-nascido pode caminhar e até mesmo correr após uma hora do nascimento.
Propõe-se que a evolução da reprodução placentária só se tornou possível por causa dos retrovírus que:
- Fazem a conexão entre a placenta e o útero num Syncytium, ou seja, uma fina camada de células com uma membrana externa compartilhada. Isto permite a passagem de oxigênio, nutrientes e de produtos indesejados, mas evita a passagem de sangue e outras células que causariam o ataque do sistema imunológico da mãe ao feto.
- Reduz a agressividade do sistema imunológico da mãe (o que é bom para o feto, mas torna a mãe mais vulnerável a infecções).
De um ponto de vista paleontólogo, os eutherianos são principalmente diferenciados por várias características de seus dentes.
Expansão dos nichos ecológicos no Mesozoico
Ainda há algo de verdadeiro no estereótipo "insetívoros pequenos e noturnos", mas descobertas recentes, principalmente na China, mostram que alguns mamaliformes e mamíferos verdadeiros eram grandes e tinham estilos de vida variados. Por exemplo:
- O Castorocauda, que viveu no Jurássico Médio, há cerca de 164 milhões de anos, tinha cerca de 43 cm de comprimento, pesava cerca de 800 g, tinha membros que eram adaptados para nadar e cavar e dentes adaptados para comer peixes.
- Os multituberculados, que sobreviveram durnate cerca de 120 milhões de anos (do Jurássico Médio, há cerca de 160 milhões de anos, ao começo do Oligoceno, há cerca de 35 milhões de anos), são frequentemente chamados de "roedores do Mesozoico", porque eles tinham incisivos de crescimento contínuo como os modernos roedores.
- O Fruitafossor, do período final do Jurássico, há cerca de 150 milhões de anos, era quase do tamanho de uma tâmia e seus dentes, membros dianteiros e dorso sugerem que ele rompia ninhos de insetos sociais para capturá-los (provavelmente cupins, porque as formigas ainda não haviam surgido).

- O Volaticotherium antiquus, da fronteira entre o final do Jurássico e o início do Cretáceo, há cerca de 140-120 milhões de anos, é o mais antigo mamífero planador conhecido e possuía uma membrana que se estendia entre seus membros, da mesma forma do moderno esquilo-voador. Isto também sugere que ele era ativo principalmente durante o dia.
- O Repenomamus, do começo do Cretáceo, há cerca de 139-128 milhões de anos, era um predador troncudo, semelhante a um texugo, que às vezes caçava pequenos dinossauros. Duas espécies foram reconhecidas, uma com mais de 1 metro de comprimento e pesando cerca de 12–14 kg, e a outra com menos de 0,5 metro de comprimento e pesando cerca de 4–6 kg.
Evolução dos principais grupos de mamíferos atuais
Há atualmente debates vigorosos entre os paleontólogos tradicionais ("caçadores de fósseis") e os filogeneticistas moleculares sobre como e quando os mamíferos verdadeiros se diversificaram, especialmente os placentários. Geralmente, os paleontólogos tradicionais datam o surgimento de um grupo particular pelo mais antigo fóssil conhecido cujas características fazem-no provavelmente ser um membro daquele grupo, enquanto os filogeneticistas sugerem que cada linhagem divergiu antes (geralmente no Cretáceo) e que os mais antigos membros de cada grupo eram anatomicamente muito similares aos primeiros membros dos outros grupos e diferiam apenas em seus genes. Estes debates se estendem à definição e à relação entre os principais grupos de placentários - a controvérsia sobre a Afrotheria é um bom exemplo.
Árvore genealógica baseada em fósseis dos mamíferos placentários
Aqui uma versão bastante simplificada de uma típica árvore genealógica baseada em fósseis, tendo como base Cladogram of Mammalia - Palaeos. Ela tenta mostrar o que existe do mais próximo atualmente de uma visão consensual, mas alguns paleontólogos têm pontos de vista bastantes diferentes, por exemplo:
- O ponto de vista mais comum é de que os placentários se originaram no hemisfério sul, mas alguns paleontólogos argumentam que eles apareceram primeiramente na Laurásia (o antigo supercontinente que continha os modernos continentes da Ásia, América do Norte e Europa).
- Os paleontólogos discordam sobre quando os primeiros placentários apareceram, com estimativas abrangendo de 20 milhões de anos antes do fim do Cretáceo até quase depois do fim do Cretáceo. E os biólogos moleculares afirmam que esta origem é muito anterior.
- A maioria dos paleontólogos sugere que os placentários deveriam ser divididos em Xenarthra e o restante, mas poucos acreditam que estes animais divergiram depois.
Por segurança, brevidade e simplicidade, o diagrama omite alguns grupos extintos para dar foco à ancestralidade dos grupos modernos bem conhecidos de placentários. O diagrama também mostra:
- A época do mais antigo fóssil conhecido em muitos grupos, visto que um dos principais debates entre os paleontólogos tradicionais e os filogeneticistas moleculares é sobre quando os vários grupos começaram a se tornar distintos.
- Os membros modernos mais conhecidos da maioria dos grupos.

Esta árvore genealógica contém algumas surpresas e quebra-cabeças. Por exemplo:
- Os parentes vivos mais próximos dos cetáceos (baleias, golfinhos e botos) são artiodátilos, animais de cascos que são quase todos absolutamente vegetarianos.
- Os morcegos são claramente parentes próximos dos primatas.
- Os parentes vivos mais próximos dos elefantes são os sirénios aquáticos, e depois destes, os mais próximos são os hiraxes, que mais se parecem com os gordos porquinhos da Índia.
- Há uma pequena correspondência entre a estrutura da família (quem é descendente de quem) e as datas dos mais antigos fósseis de cada grupo. Por exemplo, os mais antigos fósseis dos perissodáctilos (membros viventes: cavalos, rinocerontes e tapires) datam do final do Paleoceno, mas os mais antigos fósseis de seu "grupo irmão", o Tubulidentata, datam do começo do Mioceno, aproximadamente 50 milhões de anos depois. Os paleontólogos são bastante confiantes quanto a estas relações, que são baseadas em análises cladísticas, e acreditam que os fósseis dos ancestrais dos modernos aardvarks simplesmente ainda não foram encontrados.
Árvore genealógica dos mamíferos placentários de acordo com a filogenética molecular
A filogenética molecular usa características dos genes dos organismos para desenvolver árvores genealógicas da mesma forma que os paleontólogos fazem com as características dos fósseis - se genes de dois organismos são mais similares entre si que os de um terceiro organismo, os dois organismos são mais proximamente relacionados entre si que com o terceiro.
Os filogeneticistas moleculares propuseram uma árvore genealógica que é muito diferente daquela que os paleontólogos estão familiarizados. Como os paleontólogos, os filogeneticistas moleculares têm opiniões diferentes sobre vários detalhes, mas a árvore genealógica aqui apresentada é típica de acordo com os filogeneticistas moleculares. Note que o diagrama aqui mostrado omite os grupos extintos, porque não se pode extrair DNA de fósseis.

As mais significativas das muitas diferenças entre esta árvore genealógica e a familiar aos paleontólogos são:
- A divisão no mais alto nível é entre Atlantogenata e Boreoeutheria, no lugar de entre Xenarthra e o restante. Mas alguns filigeneticistas moleculares propõem uma divisão tríplice: Xenarthra, Afrotheria e Boreoeutheria.
- Afrotheria contém vários grupos que são apenas relacionados distantemente de acordo com a versão dos paleontólogos: Afroinsectiphilia ("insetívoros africanos"), Tubulidentata (aardvarks, que os paleontólogos consideram como parentes mais próximos dos ungulados de cascos ímpares que outros membros da Afrotheria), Macroscelidea (musaranhos-elefantes, normalmente considerados como próximos de coelhos e roedores). Os únicos membros da Afrotheria que os paleontólogos considerariam como parentes próximos são: Hyracoidea (hiraxes), Proboscidea (elefantes) e Sirenia (peixes-boi, dugongos).
- Os insetívoros estão divididos em três grupos: um faz parte do Afrotheria e os outros dois são sub-grupos distintos dentro do Boreoeutheria.
- Os morcegos estão mais próximos dos carnívoros e dos ungulados de cascos ímpares que dos primatas e do Dermoptera (colugos).
- Os perissodáctilos (ungulados de cascos ímpares) estão mais próximos dos carnívoros e morcegos que dos artiodáctilos (ungulados de cascos pares).
O agrupamento Afrotheria tem de certa forma justificativa geológica. Todos os membros sobreviventes do Afrotheria vivem na América do Sul ou (principalmente) na África, devido ao supercontinente Pangea se ter separado dos outros continentes há cerca de 150 milhões de anos, e depois se ter dividido em América do Sul e África, há entre 100 e 80 milhões de anos. O mais antigo mamífero euteriano conhecido é o Eomaia, de há cerca de 125 milhões de anos. Então, não seria surpreendente se os mais antigos imigrantes euterianos na África e na América do Sul ficaram isolados e se irradiaram para todos os nichos ecológicos disponíveis.
Todavia, estas proposições são controversas. Os paleontólogos naturalmente insistem que a evidência fóssil deve ter prioridade sobre as deduções de amostras de DNA de animais modernos. De maneira ainda mais surpreendente, estas novas árvores genealógicas têm sido criticadas por outros filogeneticistas moleculares, às vezes com bastante dureza:
- A taxa de mutação do DNA mitocondrial nos mamíferos varia de região para região - algumas partes raramente mudam e outras mudam radicalmente depressa e até mesmo mostram grandes variações entre indivíduos dentro das mesma espécie.
- O DNA mitocondrial mamífero modifica-se tão rapidamente que causa um problema chamado "saturação", onde as interferências aleatórias suprimem qualquer informação que talvez esteja presente. Se um pedaço particular de DNA mitocondrial se modifica aleatoriamente durante uns poucos milhões de anos, ele terá mudado várias vezes nos últimos 60 a 75 milhões de anos, quando os principais grupos de mamíferos placentários divergiram.
Evolução placentária: quando começou?
Estudos filogenéticos moleculares recentes sugerem que a maioria das ordens placentárias divergiram há cerca de 100 a 80 milhões de anos, mas as famílias modernas surgiram pela primeira vez no final do Eoceno e no começo do Mioceno.
Alguns paleontólogos objetam que nenhum fóssil placentário foi encontrado antes do fim do Cretáceo - por exemplo, o Maelestes gobiensis, de há cerca de 75 milhões de anos, é um euteriano, mas não um placentário verdadeiro.
Os fósseis dos mais antigos membros da maioria dos modernos grupos datam do Paleoceno, alguns são posteriores e pouquíssimos datam do Cretáceo, de antes da extinção dos dinossauros. Mas alguns paleontólogos, influenciados pelos estudos filogenéticos moleculares, usaram métodos estatísticos para extrapolar mais para o passado fósseis dos grupos modernos e concluíram que os primatas surgiram no final do Cretáceo.
Evolução das características dos mamíferos
Mandíbulas e ouvidos médios
O Hadrocodium, cujos fósseis datam do começo do Jurássico, fornece a primeira evidência clara de articulação da mandíbula e ouvido médio completamente mamíferos, onde a articulação da mandíbula é formada pelos ossos dentário e esquamosal enquanto os ossos articular e quadratum se deslocaram para o ouvido médio, transformando-se nos ossos bigorna e martelo. Curiosamente, ele é normalmente classificado como um membro do grupo mamaliformes e não como um mamífero verdadeiro.
Tem sido proposto que o ouvido médio tipicamente mamífero desenvolveu-se duas vezes independentemente, nos monotremados e nos mamíferos terianos, mas essa é uma opinião contestada.
Lactação
Sugere-se que a função original da lactação era manter os ovos úmidos. O argumento é baseado em grande parte nos monotremados (mamíferos que põem ovos):
- Os monotremados não possuem mamilos, mas secretam leite por glândulas peludas em seus abdômens.
- Durante a incubação, os ovos dos monotremados são cobertos por uma substância pegajosa cuja origem é desconhecida. Antes de os ovos serem postos, eles possuem uma casca com três camadas. Depois de postos, uma quarta camada aprece, e sua composição é diferente das três originais. A substância pegajosa e a quarta camada talvez sejam produzidas por glândulas mamárias.
- Se é assim, isto talvez explique porque as glândulas das quais os monotremados secretam leite sejam peludas - é fácil espalhar umidade e outras substâncias sobre o ovo a partir de uma área ampla e peluda que de um mamilo pequeno e sem pelos.
Pêlos e pele
A primeira evidência clara de cabelos ou pêlos está nos fósseis do Castorocauda, de há 164 milhões de anos, no Jurássico Médio.
De 1955 em diante, alguns cientistas interpretaram que as fendas (passagens) nos maxilares (mandíbulas superiores) e pré-maxilares (pequenos ossos na frente dos maxilares) dos cinodontes como canais que forneciam vasos sangüíneos e nervos para bigodes sensoriais (vibrissas), e sugeriram que isto era uma evidência de cabelos ou pêlos. Mas estas aberturas não necessariamente mostram que um animal possua vibrissas - por exemplo, o moderno lagarto Tupinambis possui aberturas que são quase idênticas àquelas encontradas no cinodonte não-mamífero Thrinaxodon.
Membros eretos
A evolução dos membros eretos nos mamíferos está incompleta - monotremados atuais e fósseis possuem membros não eretos. Na realidade, alguns cientistas acham que a postura parassagital (membros eretos) é uma sinapomorfia (característica diferenciada) do Boreosphenida, um grupo que contém o Theria e, portanto, inclui o último ancestral comum dos marsupiais e placentários modernos - e dessa forma, mostra que todos os primeiros mamíferos possuiam membros não eretos.
O Sinodelphys (o mais antigo marsupial conhecido) e o Eomaia (o mais antigo placentário conhecido) viveram há cerca de 125 milhões de anos, de modo que os membros eretos devem ter evoluído antes deles.
Sangue quente
"Sangue quente é um termo complexo e particularmente ambíguo, porque ele inclui alguns ou todos os termos abaixo:
- Endotermia, ou seja, a capacidade de gerar calor internamente e não através de comportamentos tais como aquecer-se ao sol ou de atividade muscular.
- Homeotermia, ou seja, a manutenção bastante constante da temperatura corporal.
- Taquimetabolismo, ou seja, um tipo de termorregulação usada por criaturas que mantêm um alto metabolismo de descanso. Criaturas taquimetabólicas estão, essencialmente, "ligadas" o tempo todo. De qualquer forma, seus metabolismos de descanso são ainda muitas vezes mais lentos que seus metabolismos ativos. A diferença não é frequentemente tão grande como a vista nas criaturas de sangue frio. As criaturas taquimetabólicas têm grande dificuldade de lidar com uma escassez de comida.
Visto que nós não podemos saber muito sobre os mecanismos internos das criaturas extintas, a maioria das discussões se concentra na homeotermia e no taquimetabolismo.
Os monotremados modernos têm uma temperatura corporal baixa e taxa metabólica mais variável que os marsupiais e placentários. Então, a questão principal é quando um metabolismo monotremado evoluiu nos mamíferos. As evidências encontradas até agora sugerem que cinodontes triássicos devem ter tido taxas metabólicas bastante elevadas, mas isto não é conclusivo.
Concha nasal respiratória
Os mamíferos modernos têm conchas nasais respiratórias, estruturas convolutas de osso fino na cavidade nasal. Estas estão alinhadas com as mucosas que aquecem e umedecem o ar inalado e extraem calor e umidade do ar exalado. Um animal com conchas nasais respiratórias pode manter uma alta taxa de respiração sem o perigo de secar os seus pulmões, e assim poder ter um metabolismo acelerado. Por azar, estes ossos são muito delicados e dessa forma ainda não foram encontrados em fósseis. Mas, sulcos rudimentares como aqueles que sustentam as conchas nasais respiratórias foram encontradas em terápsidas triássicos como o Thrinaxodon e o Diademodon, o que sugere que eles talvez tivessem taxas metabólicas bastante elevadas.
Palato secundário ósseo
Os mamíferos possuem um palato secundário ósseo que separa a passagem respiratória da boca, permitindo que eles comam e respirem ao mesmo tempo. Palatos secundários ósseos foram encontrados nos cinodontes mais avançados e têm sido usados como evidência de taxas metabólicas elevadas. Mas alguns vertebrados de sangue frio possuem palatos secundários ósseos (crocodilianos e alguns lagartos), enquanto as aves, que são animais de sangue quente, não os possuem.
Diafragma
Um diafragma muscular ajuda os mamíferos a respirarem, especialmente durante atividades enérgicas. Para um diafragma funcionar, as costelas não devem restringir o abdômen, de forma que a expansão do tórax pode ser compensada pela redução no volume do abdômen e vice-versa. Os cinodontes mais avançados tinham caixas torácicas bastante semelhantes às dos mamíferos, com costelas lombares muito reduzidas. Isto sugere que estes animais tinham diafragmas, eram capazes de atividades extenuantes por períodos bastante longos e então tinham altas taxas metabólicas. Por outro lado, estas caixas torácicas semelhantes às dos mamíferos podem ter se desenvolvido para aumentar a agilidade. Mas este passo de terapsídeos mais avançados foi "como um carro de mão", com os membros traseiros fornecendo todo o impulso enquanto os membros dianteiros apenas guiando o animal, em outras palavras, estes terapsídeos avançados não eram tão ágeis quanto os mamíferos modernos ou os primeiros dinossauros. Então, a ideia de que a principal função destas cavidades torácicas semelhantes às dos mamíferos era de aumentar a agilidade é duvidável.
Postura dos membros
Os terapsídeos tinham os membros dianteiros estendidos e os traseiros semi-eretos. Isto sugere que a restrição de Carrier teria tornado para eles particularmente difícil se mover e respirar ao mesmo tempo, mas não tão difícil para animais como como os lagartos que têm membros completamente estendidos. Mas os cinodontes (terapsídeos avançados) tinham placas nas costelas que endureciam a cavidade torácica e assim possivelmente reduziam a flexão lateral do tronco enquanto se moviam, o que teria tornado um pouco mais fácil para eles respirar enquanto se moviam. Estes fatos sugerem que os terapsídeos avançados eram significativamente menos ativos que os mamíferos modernos de tamanho similar e dessa forma talvez tivessem metabolismos lentos.
Isolamento térmico (cabelos e pêlos)
O isolamento térmico é o caminho menos custoso para se manter a temperatura do corpo constante. Assim, possuir cabelos ou pêlos seria uma boa evidência de homeotermia, mas não tão forte quanto a evidência de uma alta taxa metabólica .
Vimos que: a primeira evidência clara de cabelos ou pêlos está em fósseis do Castorocauda, de há 164 milhões de anos, no Jurássico Médio; os argumentos de que os terapsideos avançados tinham pêlos não são convincentes.
Bibliografia
- Robert L. Carroll, Vertebrate Paleontology and Evolution, W. H. Freeman and Company, New York, 1988 ISBN 0-7167-1822-7. Do Capítulo XVII até o XXI.
- Nicholas Hotton III, Paul D. MacLean, Jan J. Roth, and E. Carol Roth, editors, The Ecology and Biology of Mammal-like Reptiles, Smithsonian Institution Press, Washington and London, 1986 ISBN 0-87474-524-1
- T. S. Kemp, The Origin and Evolution of Mammals, Oxford University Press, New York, 2005 ISBN 0-19-850760-7
- Zofia Kielan-Jaworowska, Richard L. Cifelli, and Zhe-Xi Luo, Mammals from the Age of Dinosaurs: Origins, Evolution, and Structure, Columbia University Press, New York, 2004 ISBN 0-231-11918-6. Cobertura abrangente dos primeiros mamíferos até a época da extinção em massa K-T.
- Zhe-Xi Luo, "Transformation and diversification in early mammal evolution", Nature volume 450 number 7172 (13 December 2007) pages 1011-1019. doi:10.1038/nature06277. Um artigo de perquisa com 98 referências para a literatura científica.
Ligações externas
- The Cynodontia trata vários aspectos da evolução de cinodontes em mamíferos, com abundância de referências.