
Codificação neural é um campo relacionado à neurociência que se preocupa em caracterizar as relações entre o estímulo e o indivíduo ou juntar as respostas neuronais e a relação entre a atividade elétrica dos neurônios no conjunto. Baseada na teoria de que a informação sensorial e outras estão representadas no cérebro por redes neurais biológicas, pensa-se que os neurônios podem codificar ambas as informações digitais e analógicas.
Resumo
Os neurônios são notáveis entre as células do corpo em sua habilidade para propagar sinais rapidamente por longas distâncias. Eles fazem isso gerando pulsos elétricos característicos chamados de potenciais de ação: pulos com voltagem que podem viajar pelas fibras nervosas. Neurônios sensoriais mudam suas atividades ao disparar sequências de potenciais de ação em vários padrões temporais, com a presença de estímulo sensorial externo, como luz, som, sabor, cheiro e tato. Sabe-se que a informação sobre o estímulo está codificada neste padrão de potenciais de ação e é transmitida para e ao redor do cérebro.
A pesar de que os potenciais de ação podem variar em duração, amplitude e forma, eles são tipicamente tratados como eventos generalizadamente idênticos nos estudos sobre codificação neural. Se a breve duração de um potencial de ação (aproximadamente 1ms) é ignorada, uma sequência de potencial de ação, ou trem de pulsos nervosos, pode ser caracterizada simplesmente por uma série de eventos ponto de tudo ou nada no tempo. A duração dos intervalos inter-spikes (ISI) entre dois pulsos nervosos sucessivos em um trem de pulsos nervosos varia com frequência, aparentemente aleatoriamente. O estudo da codificarão neural envolve medir e caracterizar como os atributos de estímulos como luz ou intensidade sonora, ou ações motoras, como a direção do movimento do braço, são representados pelos potenciais de ação dos neurônios ou pulsos nervosos. A fim de descrever e analisar o disparo dos neurônios, métodos estatísticos, métodos da teoria da probabilidade e processos pontuais estocásticos tem sido amplamente aplicados.
Com o desenvolvimento de tecnologias de grande escala de registro e decodificação neural, pesquisadores tem começado a entender o código neural e já deram a primeira olhadela em tempo real ao código neural enquanto a memória é formada e lembrada no hipocampo, uma região cerebral conhecida como a parte central para a formação da memória. Neurocientistas iniciaram vários projetos de grande escala para decodificar o cérebro.
Codificando e decodificando
O link entre estimulo e resposta pode ser estudado desde dois pontos de vista opostos. Codificação neural se refere ao mapa do estimulo à resposta. O foco principal é entender como os neurônios respondem a uma variedade de estímulos, e construir modelos que tentem prever respostas para outros estímulos. Decodificação neural se refere ao mapa inverso, da resposta para o estímulo, e o desafio é reconstruir um estímulo, ou certos aspectos daquele estímulo, a partir das sequências de pulsos nervosos que ele evoca.
Esquemas de codificação
Uma sequência, ou "trem", de pulsos nervosos pode conter informação baseada em diferentes esquemas de codificação. Em neurônios motores, por exemplo, a força com a qual um músculo inervado se flexiona depende apenas da "taxa de disparo", o número médio de pulsos nervosos por unidade de tempo ( uma "taxa de código"). Por outro lado, um "código temporal" complexo é baseado no preciso tempo de um único pulso nervoso. Eles podem estar presos a um estímulo externo, como no sistema visual ou auditivo, ou ser gerado intrinsicamente pelo circuito neural.
Se os neurônio usam codificação por ritmo ou temporal é um tema de debate intenso dentre a comunidade da neurociência, mesmo não havendo uma definição clara sobre o que esses termos significam. Em uma teoria, chamada "neuro-eletrodinâmica", os esquemas de codificação seguintes são todos considerados como epifenômenos, substituídos, ao contrário, por mudanças moleculares refletindo a distribuição espacial de campos elétricos dentre os neurônios como um resultado do amplo espectro eletromagnético dos potenciais de ação, e manifestado como informação como diretividade de pulsos neurais.
Codificação por ritmo
O modelo de codificação por ritmo da comunicação entre os disparos dos neurônios declara que a medida que a intensidade do estímulo aumenta, a freqüência ou ritmo dos potenciais de ação, ou "disparo dos pulsos nervosos", aumenta. A codificação por ritmo é às vezes chamada de codificação por freqüência.
A codificação por ritmo é um esquema de codificação tradicional, assumindo que a maioria, se não toda, a informação sobre o estímulo é contida no ritmo de disparo do neurônio. Porque a sequência dos potenciais de ação gerada por um dado estímulo varia de experimento a experimento, as respostas neurais são tipicamente tratadas probabilística ou estatisticamente. Elas podem ser caracterizadas pelo ritmo dos disparos, ao invés de por sequências de pulsos nervosos específicas. Na maior parte dos sistemas sensoriais, o ritmo dos disparos aumenta, geralmente não linearmente, com o aumento da intensidade do estímulo. Qualquer possível informação codificada na estrutura temporal do trem de pulsos nervosos é ignorada. Consequentemente, a codificação por ritmo é ineficiente mas altamente robusta em relação ao "ruído" do ISI.
Durante a codificação por ritmo, calcular o ritmo dos disparos de forma precisa é muito importante. De fato, o termo "ritmo de disparo" tem algumas definições diferentes, as quais se referem a diferentes procedimentos para tirar a média, como a média tirada a partir do tempo ou a média tirada a partir de várias repetições do experimento. Para a codificação por ritmo, o ato de aprender esta baseado no peso das modificações sinápticas dependentes de atividade.
A codificação por ritmo foi originalmente mostrada por ED Adrian e Y Zotterman em 1926. Neste simples experimento diferentes pesos eram dependurados de um músculo. A medida que o peso do estímulo aumentava, o número de pulsos nervosos registrados desde os nervos sensoriais que inervavam o músculo também aumentavam. A partir destes experimentos originais, Adrian e Zotterman concluíram que os potenciais de ação eram eventos unitários, e que a freqüência dos eventos, não a magnitude de eventos individuais, era a base para a maior parte da comunicação inter-neural.
Nas décadas seguintes, a medição dos ritmos de disparo se tornou uma ferramenta padrão para descrever as propriedades de todos os tipos de neurônios sensoriais ou corticais, parcialmente devido à relativa facilidade de tomar as medidas experimentalmente. No entanto, esta abordagem negligencia toda a informação possivelmente contida no exato momento do pulso nervoso. Durante anos mais recentes, mais e mais evidencias experimentais têm sugerido que um conceito direto de ritmo de disparo baseado na média temporal pode ser muito simples para descrever a atividade cerebral.
Ritmo por contagem de pulsos nervosos
O ritmo por contagem de pulsos nervosos, também referido como média temporal, é obtido pela contagem do número de pulsos nervosos que aparecem durante um experimento divididos pela duração do experimento. A duração D da janela temporal é estipulada pelo investigador e depende do tipo de neurônio do qual se registra e do estímulo. Na prática, para conseguir médias sensatas, vários pulsos nervosos deveriam ocorrem durante a janela temporal. Valores típicos são D=100 ms ou D=500 ms, mas a duração também pode ser mais longa ou curta.
O ritmo por contagem de pulsos nervosos pode ser determinado desde um único experimento, mas pelo custo de perder toda a resolução temporal sobre as variações na resposta neural durante o curso do experimento. A média temporal pode servir bem em casos onde o estímulo é constante ou de variação lenta e não requer uma reação rápida do organismo- e esta é a situação normalmente encontrada nos protocolos experimentais. Inputs do mundo real, no entanto, quase nunca são estacionários, mas com freqüência mudam em uma escala temporal rápida. Por exemplo, mesmo quando observando uma imagem estática, os humanos realizam movimentos sacádicos, mudanças rápidas na direção da visão. A imagem projetada nos fotorreceptores da retina muda, então, a cada cem milissegundos, aproximadamente.
A pesar de suas fraquezas, o conceito de ritmo por contagem de pulsos nervosos é amplamente usado não apenas em experimentos, mas também em modelos de redes neurais. O conceito levou à ideia de que um neurônio transforma informação sobre uma única variável de entrada (a força do estímulo) em uma única saída contínua e variável (o ritmo dos disparos).
Há um crescente corpo de evidencias de que nos neurônios de Purkinje, ao menos, a informação não é codificada simplesmente nos disparos mas também no momento e duração dos "não-disparos", períodos de silêncio.
Ritmos de disparo dependente do tempo
O ritmo de disparo dependente do tempo é definido como o número médio de pulsos nervosos (média feita durante os experimentos) aparecendo durante um curto intervalo entre tempos t e t+Δt, divididos pela duração do intervalo. Ele funciona tanto para estímulos estacionários quanto para estímulos dependentes de tempo. Para medir experimentalmente o ritmo de disparo dependente de tempo, o investigador registra um neurônio enquanto estimulado por alguma sequência de entrada. A mesma sequência de estimulação é repetida várias vezes e a resposta neural é relatada em uma histograma de tempos peri-estímulo (PSTH). O tempo t é medido com respeito ao começo da sequência de estimulação. O Δt deve ser grande o suficiente (tipicamente entre um ou alguns poucos milissegundos) para que haja número suficiente de pulsos nervosos dentro do intervalo para obter uma estimativa da média confiável. O número de ocorrências de pulsos nervosos nK(t;t+Δt) somado sobre todas as repetições do experimento divido pelo número K de repetições é a medida da atividade típica de um neurônio entre o tempo e e t+Δt. Uma divisão adicional pela duração do intervalo Δt contém o ritmo de disparo dependente de tempo r(t) do neurônio, o qual é equivalente à densidade dos pulsos nervosos do PSTH.
Para um Δt suficientemente pequeno, r(t)Δt é o número médio de pulsos nervosos ocorrendo entre os tempos t e t+Δt em múltiplos experimentos. Se Δt é pequeno, nunca haverá mais do que um pulso nervoso dentro do intervalo t e t+Δt em qualquer dos experimentos. Isso significa que r(t)Δt é também a fração dos experimentos nos quais um pulso nervoso ocorreu entre esses tempos. Equivalentemente, r(t)Δt é a probabilidade de que um pulso nervoso ocorrerá durante esse intervalo de tempo.
Como um procedimento experimental, a medida do ritmo de disparo dependente do tempo é um método útil para avaliar a atividade neural, em particular no cado do estímulo dependente do tempo. O problema óbvio com esta abordagem é que ela não pode ser o esquema de codificação usado pelo neurônio no cérebro. Neurônios não podem esperar que os estímulos se apresentem repetidamente e na mesma maneira antes de gerar uma resposta.
No entanto, a medida experimental do ritmo de disparo dependente do tempo pode fazer sentido se há uma grande população de neurônios independentes que recebem o mesmo estímulo. Ao invés de registrar uma população de N neurônios em uma única corrida, é experimentalmente mais fácil registrar um único neurônio e tomar a média sobre N corridas repetidas. Então, a codificação de ritmo de disparo dependente do tempo se apoia nas suposições de que sempre há populações de neurônios.
Codificação temporal
Quando flutuações precisas no tempo ou ritmo de disparo de alta frequência dos pulsos nervosos são encontradas carregando informação, o código neural é com frequência identificado como um código temporal. Um número de estudos encontraram que a resolução temporal do código neural está em uma escala temporal de milissegundos, indicando que o tempo preciso dos pulsos nervosos é um elemento significativo na codificação neural.
Os neurônios exibem flutuações de alta freqüência nos ritmos de disparo, o que pode ser tanto ruído quanto informação. Modelos de codificação por ritmo sugerem que essas irregularidades são ruídos, enquanto que modelos de codificação temporal sugerem que eles codificam informação. Se o sistema nervoso fizesse uso apenas de códigos de ritmo para transmitir informação, um ritmo de disparo mais consistente, regular, seria evolucionariamente vantajoso, e os neurônios teriam usado este código ao invés de outras opções menos robustas. A codificação temporal fornece uma explicação alternativa para o "ruído", sugerindo que ele na verdade codifica informação e afeta o processamento neural. Para modelar esta ideia, símbolos binários podem ser usados para marcar os pulsos nervosos: 1 para pulso e 0 para não pulso. A codificação temporal permite que a sequência 000111000111 signifique algo diferente de 001100110011, mesmo que a média de disparos seja a mesma para ambas as sequências, a 6 pulsos/10 ms. Até recentemente, cientistas haviam posto maior ênfase na codificação por ritmo como uma explicação para os padrões de potencial pós-sinápticos. No entanto, as funções do cérebro são mais temporariamente precisas do que aquilo que o uso de codificação por ritmo parece permitir. Em outras palavras, informação essencial poderia ser perdida devido à inabilidade do código por ritmo de capturar toda a informações disponível no trem de pulsos nervosos. Além disso, respostas são suficientemente diferentes entre estímulos similares (mas não idênticos) como para sugerir que os padrões distintos dos pulsos nervosos contém um volume maior de informação do que aquilo que é possível incluir em um código por ritmo.
Os códigos temporais fazem uso das características da atividade dos pulsos nervosos que não podem ser descritas pelo ritmo de disparo. Por exemplo, o tempo para o primeiro pulso nervosos depois do estimulo inicial, características baseadas no segundo e maior momento estatístico da distribuição de probabilidade do ISI, aleatoriedade do pulso nervoso, ou precisamente grupos com padrões temporais são candidatos para códigos temporais. Como não há nenhuma referência temporal absoluta no sistema nervoso, a informação é carregada ou em termos de tempo relativo dos pulsos nervosos em uma população de neurônios ou com respeito a uma oscilação cerebral em andamento.
A estrutura temporal de um trem de pulso nervoso ou um ritmo de disparo evocado por um estimulo é determinado ambos pela dinâmica do estímulo e pela natureza do processo de codificação neural. Estímulos que mudam rapidamente tendem a gerar pulsos de tempo preciso e ritmos de disparos de rápida mudança sem importar qual estratégia de codificação neural está sendo usada. Codificação temporal se refere à precisão temporal na resposta que não surge apenas da dinâmica do estímulo, mas que não obstante se relaciona à propriedades do estímulo. A interação entre o estímulo e as dinâmicas de codificação tornam a identificação de um código temporal difícil.
Na codificação temporal, o aprendizado pode ser explicado por modificações sinápticas dependentes de atividade atrasadas. As modificações podem elas mesmas depender não apenas dos ritmos do pulso (codificação por ritmo) mas também dos padrões temporais do pulso (codificação temporal); e.x. pode ser um caso especial de plasticidade de pulso dependente de tempo.
A questão da codificação temporal é distinta e independente da questão da codificação de pulso independente. Se cada pulso é independente de todos os outros pulsos no trem, o caráter temporal do código neural é determinado pelo comportamento do ritmo de disparo dependente de tempo r(t). Se r(t) varia lentamente com o tempo, o código é tipicamente chamado de código por ritmo, e se ele varia rapidamente, o código é chamado de temporal.
Codificação temporal em sistemas sensoriais
Para estímulos muito breves, o ritmo máximo de disparo de um neurônio pode não ser rápido o suficiente para produzir mais de um único pulso. Devido à densidade de informação sobre o estimulo abreviado contido neste único pulso, pareceria que o tempo do pulso em si teria de carregar mais informação do que simplesmente a freqüência média dos potenciais de ação sobre um dado período de tempo. Este modelo é especialmente importante para a localização sonora, a qual ocorre no cérebro em ordem de milissegundos. O cérebro deve obter uma quantidade imensa de informação baseado em uma relativamente curta resposta neural. Além disso, se ritmos baixos de disparo na ordem de dez pulsos por segundo devem ser distinguidos de ritmos de codificação arbitrariamente próximos para estímulos diferentes, então um neurônio que tente discriminar esses dois estímulos pode precisar esperar por um segundo ou mais para acul=mular informação suficiente. Isto não é consistente com numerosos organismos que são capazes de discriminar entre estímulos no prazo de milissegundos, sugerindo que um código de ritmo não é o único modelo funcionando.
Para justificar a rapida codificação de estímulos visuais, foi sugerido que os neurônios da retina codificam informação visual no tempo de latência entre o estímulo inicial e o potencial de ação, também chamado de latência para o primeiro pulso. Este tipo de codificação temporal tem também sido mostrada no sistema auditivo e somato-sensitivo. A principal inconveniência de tal esquema de codificação é sua sensibilidade à flutuações neurais intrínsecas. No córtex visual primário de macacas, descobriu-se que o tempo do primeiro pulso relativo ao começo do estímulo prove mais informação do que o intervalo entre pulsos. No entanto, o intervalo entre pulsos poderia ser usado para codificar informação adicional, que é especialmente importante quando o ritmo do pulso alcança seu limite, como em situações de alto contraste. Por essa razão, a codificação temporal pode ter parte na codificação de bordas definidas, ao invés de transições graduais.
O sistema gustativo dos mamíferos é útil para estudar codificação temporal por causa de seus estímulos razoavelmente distintos e as respostas facilmente discerníveis do organismo. Informação temporariamente codificada pode ajudar um organismo a discriminar entre diferentes gostos da mesma categoria (doce, amargo, azedo, salgado, umami) que obtém respostas muito similares em termos de contagem de pulsos. O componente temporal do padrão obtido por cada gosto pode ser usado para determinar sua identidade (e.x. a diferença entre dois gostos amargos, como a quinina e o denatônio). Desta forma, ambas a codificação por ritmo e codificação temporal podem ser usadas no sistema gustativo-o ritmo para tipos de gosto básicos e o temporal para diferenciações mais específicas. Pesquisas sobre o sistema gustativo dos mamíferos mostraram que há uma abundância de informação presente em padrões temporais através das populações de neurônios, e essa informação é diferente daquele que é determinada pelo esquema de codificação por ritmo. Grupos de neurônios podem sincronizar em resposta a um estímulo. Em estudos lidando com a porção frontal do córtex cerebral em primatas, padrões precisos com escalas temporais curtas de apenas alguns poucos milissegundos de duração foram encontrados por entre pequenas populações de neurônios que se correlacionavam com certos comportamentos de processamento de informação. No entanto, pouca informação pode ser determinada a partir dos padrões; uma possível teoria é que eles representavam o processamento de ordem mais alta que acontecia no cérebro.
Como com o sistema visual, nas células mitrais/em tufo no bulbo olfatório dos ratos, a latência do primeiro pulso relativo ao começo da ação de cheirar pareceu modificar muita da informação sobre o odor. A estratégia de usar a latência dos pulsos permite uma rápida identificação para a reação a um cheiro. Além disso, algumas células mitrais/em tufo tem padrões de disparo específicos para dados aromas. Este tipo de informação extra poderia ajudar no reconhecimento de um certo cheiro, mas não é completamente necessário, uma vez que a média da contagem de pulsos enquanto os animais cheiravam foi também um bom identificador. Na mesma linha que este experimento, experimentos feitos com o sistema olfativo de coelhos mostrou padrões distintos que se correlacionavam com diferentes subconjuntos de aromas, um resultado similar foi obtido em experimentos com o sistema olfativo de gafanhotos.
Aplicações da codificação temporal
A especificidade da codificação temporal requer tecnologia altamente refinada para medir dados experimentais informativos e confiáveis. Progressos feitos na optogenética permitem à neurologistas controlar pulsos em neurônios individuais, oferecendo resolução elétrica e espacial à células individuais. Por exemplo, a luz azul faz com que o canal rodopsina se abra, despolarizando a célula e produzindo um pulso. Quando a luz azul não é sentida pela célula, o canal se fecha, e o neurônio para de pulsar. O padrão dos pulsos combina com o padrão do estímulo da luz azul. Inserindo sequências genéticas do canal rodopsina no DNA de um rato, pesquisadores podem controlar os pulsos e, dessa forma, certos comportamentos do rato (e.x. fazer o rato girar à esquerda). Pesquisadores, através da optogenética, tem as ferramentas para afetar diferentes códigos temporais em um neurônio enquanto mantém a mesma média de disparos e, por consequência, podem testar se a codificação temporal ocorre ou não em circuitos neurais específicos.
A tecnologia de optogenética também tem o potencial de possibilitar a correção de anormalidades nos pulsos na raiz de vários distúrbios neurológicos e psicológicos. Se os neurônio de fato codificam informação em pulsos de padrões temporais específicos, sinais chaves poderiam se perder ao tentar desvendar o código olhando apenas para a média dos disparos. Entender qualquer aspecto temporariamente codificado do código neural e replicar essas sequências em neurônios poderia permitir um maior controle e tratamento de distúrbios neurológicos com depressão, esquizofrenia e Parkinson. A regulação dos intervalos entre os pulsos em células individuais controla a atividade cerebral e forma mais precisa do que a adição de agentes farmacológicos intravenosamente.
Código por disparo de fase
O código por disparo de fase é um esquema de codificação neural que combina a contagem de pulsos com uma referência temporal baseada em oscilações. Este tipo de código leva em conta uma etiqueta para o tempo de cada pulso de acordo com uma referência de tempo baseada na fase de oscilações locais em andamento em frequências baixas ou altas. Uma característica dede código é que os neurônios se aderem a uma ordem preferida de pulsos, resultando em uma sequência de disparos.
Foi mostrado que os neurônios em algumas áreas sensoriais do córtex codificam ricos estímulos naturais em termos dos tempos do pulso relativo à fase das flutuações da rede em andamento, ao invés de apenas em termos da contagem dos pulsos. As oscilações refletem os sinais do potencial de campo local. Com freqüência ele é categorizado como um código temporal a pesar de que a etiqueta de tempo usada para os pulsos seja grosseira. Isto é, quatro valores discretos para a fase são suficientes para representar toda a informação neste tipo de código com respeito à fase das oscilações em baixa freqüência. O código por disparo de fase está imprecisamente baseado no fenômeno de procissão de fase observado nas células de lugar do hipocampo.
O código de fase tem se mostrado envolvido no córtex visual, também em oscilações de alta freqüência. Dentro de um ciclo de oscilação gama, cada neurônio tem seu próprio tempo relativo de disparo. Como resultado, uma população inteira de neurônios gera uma sequência de disparo que tem duração de até aproximadamente 15 ms.
Codificação de população
A codificação de população é um método para representar estímulos usando as atividades conjuntas de um número de neurônios. Na codificação de população, cada neurônio tem uma distribuição de respostas sobre algum conjunto de inputs, e as respostas de vários neurônios pode ser combinada para determinar algum valor sobre os inputs.
Desde o ponto de vista teórico, a codificação de população é um dos poucos problemas matemáticos bem formulados da neurociência. Ele compreende as características essenciais da codificação neural e ainda assim é simples o suficiente para a análise teórica. Estudos experimentais tem revelado que este paradigma de codificação é amplamente usado nas áreas motoras e sensoriais do cérebro. Por exemplo, na área visual do temporal médio (TM), os neurônios estão em sintonia com a direção em movimento. Em resposta a um objeto em movimento em uma direção particular, vários neurônio do TM disparam com um padrão de atividade de som corrompido e em formato de sino através da população.
A direção em movimento do objeto é recuperada a partir da atividade da população, para ser imune da flutuação existente no sinal de um único neurônio. Em um clássico exemplo no córtex motor primário, Apostolos Georgopoulos e colegas treinaram macacos a mover um joystick em direção a um objetivo iluminado. Eles descobriram que um único neurônio dispararia para múltiplas direções. No entanto, dispararia mais rápido para uma direção e mais devagar para outra, dependendo do quão perto o objetivo estava da direção "preferida" do neurônio.
Kenneth Johnson foi quem originalmente derivou que se cada neurônio representa o movimento em sua direção preferida, e a soma do vetor de todos os neurônios é calculada (cada neurônio tem uma taxa de disparo e uma direção preferida), a soma aponta na direção do movimento. Desta maneira, a população de neurônios codifica o sinal do movimento. Este código de população em particular é referido como codificação de vetor de população. Este estudo em particular dividiu o campo da fisiologia motora entre o grupo do "neurônio motor superior" do Evart, que seguiu a hipótese de que os neurônios do córtex motor contribuíram para o controle de músculos específicos, e o grupo de Georgopoulos, que estuda a representação das direções do movimento no córtex.
O laboratório de codificação neural da Universidade de Johns Hopkins liderado por Murray Sachs e Eric Young desenvolveu códigos de população lugar-tempo, chamado de código Resposta-Média-Localizada-Sincronizada (ALSR), do inglês "Averaged-Localized-Synchronized-Response", para a representação neural de estímulos acústicos auditivos. Isto explora ambos o lugar ou sincronização dentro do nervo auditivo, assim como o fechamento de fase dentro de cada fibra nervosa do nervo auditivo. A primeira representação ALSR foram vogais estáticas; As representações do ALSR da altura do som e frequências em estados de estímulos não estáticos e complexos foi demostrado para entonação vocal e representações das sílabas de consonantes-vogais. A vantagem de tais representações é que as características globais tais como entonação ou perfis de transição podem ser representadas como características globais através do nervo inteiro via ambas as codificações de taxa e lugar simultaneamente.
A codificação de população tem um número de outras vantagens também, incluindo a redução de incertezas devido à variabilidade neuronal e a habilidade para representar um número de atributos de estímulos diferentes simultaneamente. A codificação de população é também muito mais rápida que a codificação por ritmo e pode refletir mudanças nas condições do estimulo quase instantaneamente. Neurônios individuais em tais populações tem tipicamente seletividades diferentes mas sobrepostas, de forma de vários neurônios, mas não necessariamente todos, respondem a um dado estímulo.
Tipicamente, uma função de codificação tem um valor pico de forma que a atividade do neurônio é maior se o valor perceptivo é próximo do valor pico, e se torna reduzido de acordo com os valores menos próximos do valor pico.
Segue-se que o valor de fato percebido pode ser reconstruído desde o padrão total da atividade de um conjunto de neurônio. O vetor de codificação Johnson/Georgopoulos é um exemplo de média simples. Uma técnica matemática mais sofisticada para realizar tais reconstruções é o método de máxima verossimilhança baseada em uma distribuição multivariável de respostas neuronais. Esses modelos podem assumir independência, correlações de segunda ordem, ou até mesmo dependências mais detalhadas como os modelos de entropia máxima de maior ordem ou cópulas.
Código de correlação
O modelo de codificação por correlação dos disparos neuronais alega que correlações entre potenciais de ação, ou "pulsos", dentro de um trem de pulso pode carregar informação adicional para além dos simples tempo dos pulsos. Trabalhos anteriores sugeriam que a correlação entre trens de pulsos só podiam ser reduzidos, e nunca aumentar, o total de informação mútua presente nos dois trens de pulsos sobre uma característica do estímulo. No entanto, isto foi mais tarde demostrado como incorreto. A estrutura de correlação pode aumentar o conteúdo de informação se as correlações de ruído e sinal são se sinal oposto. As correlações também podem carregar informação que não está presente na média de disparos de dois pares de neurônios. Um bom exemplo disto existe no córtex auditivo do sagui anestesiado por pentobarbital, no qual um tom puro causa um aumento no número de pulsos correlacionados, mas não um aumento na média de disparos dos pares de neurônios.
Codificação de pulso independente
O modelo de codificação de pulso independente dos disparos neurais alega que cada potencial de ação individual, ou "pulso", é independente de cada outro pulso dentro do trem de pulso.
Codificação por posição
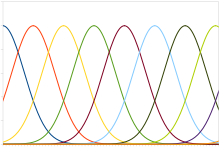
Um código de posição típico envolve neurônios com uma curva sincronizada Gaussiana, cujas médias pode variar linearmente com a intensidade do estímulo, significando que o neurônio responde mais fortemente (em termos de pulsos por segundo) a um estímulo próxima da média. A intensidade de fato poderia ser recuperada como o nível do estímulo correspondente à média do neurônio com a maior resposta. No entanto, o ruído inerente nas respostas neurais faz com que a estimação da máxima verossimilhança seja mais precisa.
Este tipo de código é usado para codificar variáveis continuas como posição conjunta, posição do olho, cor, ou freqüência sonora. Qualquer neurônio individual é muito ruidoso para codificar fielmente a variável usando a codificação por ritmo, mas uma população inteira assegura uma maior fidelidade e precisão. Para uma população de curvas tonais unimodais, ex. com um único pico, a precisão tipicamente escala linearmente com o número de neurônios. Por isso, para metade da precisão, metade dos neurônios é necessária. Em contraste, quando as curvas tonais tem múltiplos picos, como nas células de rede que representam o espaço, a precisão da população pode escalar exponencialmente com o número de neurônios. Isto reduz imensamente o número de neurônios necessários para a mesma precisão.
Codificação esparsa
A codificação esparsa é quando cada item é codificado pela forte ativação de um conjunto relativamente pequeno de neurônios. Para cada item ser codificado, um diferente subconjunto de todos os neurônio disponíveis.
Como consequência, a qualidade esparsa pode ser focada na qualidade esparsa temporal ("um número relativamente pequeno de períodos de tempo estão ativos") ou na qualidade esparsa em uma população ativa de neurônios. Neste último caso, isto pode ser definido em um período de tempo como o número de neurônios ativados relativos ao número total de neurônios da população. Isto parece ser um selo distintivo das computações neurais desde que comparada com computardes tradicionais, a informação é massivamente distribuída através dos neurônios. Um resultado importante no codificação neural de Olshausen e Field é que a codificação esparsa de imagens naturais produz filtros orientados similares à ondaletas que parecem os campos receptivos das células simples do córtex visual. A capacidade dos códigos esparsos pode ser aumentada pelo uso simultâneo da codificação temporal, como encontrado no sistema olfativo do gafanhoto.
Dado um conjunto potencialmente grade de padrões de input, algoritmos de codificação esparsa (ex. Sparse Autoencoder) tentam encontrar automaticamente um pequeno número de padrões representativos que, quando combinado na proporções certas, reproduz os padrões de input originais. A codificação esparsa para o input então consiste desses padrões representativos. Por exemplo, o conjunto muito grade de orações em Português pode ser codificado por um pequeno número de símbolos (ex. letras, pontuações, espaços) combinados em uma ordem particular para uma sentença particular, e dessa forma o código esparso para o Português seria esses símbolos.
Modelo linear generativo
A maior parte dos modelos de codificação esparsa estão baseados no modelos linear generativo. Neste modelo, os símbolos são combinados à moda linear para aproximar o input. Mais formalmente, dado um conjunto k-dimensional de vetores com input de números reais , o objetivo da codificação esparsa é determinar vetores bases n k-dimensionais junto com um vetor n-dimensinal esparso de pesos ou coeficientes para cada vetor input, de forma que uma combinação linear dos vetores bases com proporções dadas pelos coeficientes resulte em uma aproximação ao vetor input:




Os códigos gerados pelos algoritmos implementando um modelo generativo linear podem ser classificados como codificações com qualidade esparsa suave e dura. Eles se referem à distribuição dos coeficientes de vetores bases para inputs típicos. Uma codificação com qualidade esparsa suave tem uma distribuição similar à Gaussiana suave, mas com picos mais proeminentes que a Gaussiana, com vários valores em zero, alguns pequenos valores absolutos, menos valores absolutos maiores e muito menos valores absolutos muito grandes. Assim, vários dos vetores bases são ativos. A qualidade esparsa dura, por outro lado, indica que há vários valores de zero, nenhum ou quase nenhum valor absoluto pequeno, menos valores absolutos grandes e muito poucos valores absolutos muito grandes, assim menos dos vetores bases são ativos. Isto é atraente para uma perspectiva metabólica: menos energia é usada quando menor neurônios estão disparando.
Outra medida de codificação é se ela é criticamente completa ou supercompleta. Se o número de vetores bases n é igual ao conjunto input de k dimensionalidade, o código é dito ser criticamente completo. Neste caso, pequenas mudanças no vetor input resultam em mudanças abruptas nos coeficientes, e a codificação não é capaz de graciosamente manejar as pequenas escalas, pequenas translações ou ruído nos inputs. Se, ao contrário, o número de vetores bases é maior que a dimensionalidade do conjunto input, o código é supercompleto. Codificações supercompletas interpolam suavemente entre vetores input e são robusta sob o ruindo input. O córtex visual primário humano é estimado como supercompleto por um fator de 500, de forma que, por exemplo, um pedaço de input 14 x 14 (um espaço 196-dimensional) é codificado por cerca de 100.000 neurônios.
Evidência biológica
A codificação esparsa pode ser uma estratégia geral dos sistemas neurais para aumentar a capacidade da memória. Para se adaptar a seus ambientes, os animais devem aprender quais estímulos estão associados à recompensas ou repreensões e distinguir esses estímulos de outros similares, porém irrelevantes. Tais tarefas requerem memórias associativas especificas de alguns estímulos nas quais apenas alguns neurônios de uma população respondem a qualquer estímulo dado e cada neurônio responde à apenas alguns estímulos de todos os estímulos possíveis.
Trabalhos teóricos sobre memória esparsa e distribuída tem sugerido que o código esparso aumenta a capacidade da memória associativa ao reduzir sobreposição entre representações. Experimentalmente, representações esparsas de informação sensorial tem sido observada em vários sistemas, incluindo visão, audição, tato, e olfato. No entanto, a pesar do acúmulo de evidências da difusão do código esparso e os argumentos teóricos por sua importância, uma demonstração de que a codificação esparsa melhora a especificidade do estímulo da memória associativa esteve em falta até recentemente.
Algum progresso foi feito em 2014 pelo laboratório de Gero Miesenböck na Universidade de Oxford ao analisar o sistema olfativo da Drosophila. Na Drosophila, a codificação esparsa do odor feita pelas células Kenyon do corpo de cogumelo é pensada como geradora de um alto número de locais precisamente localizáveis para o armazenamento de memória de odores específicos. Lin et al. demostraram que a esparsidão é controlada por um circuito de feedback que navega entre as células Kenyon e o neurônio GABAérgico anterior lateralmente combinado (ALC). A ativação sistemática e bloqueio de cada perna deste circuito de feedback mostra que as células Kenyon ativam o ALC e o ALC inibe as células Kenyon. Interromper o loop da célula Kenyon e o ALC diminui a esparsidão das respostas ao odor da célula Kenyon, aumenta as correlações inter-odor, e impede que as moscas aprendam à discriminar odores similares, mas não não-similares. Estes resultados sugerem que a inibição do feedback suprime a atividade da célula Kenyon para manter a codificação do odor esparsa e não relacionada e, desta forma, as memórias de odores específicos.
Ver também
Notas
- Este artigo foi inicialmente traduzido, total ou parcialmente, do artigo da Wikipédia em inglês cujo título é «Neural coding».