
- Para informações sobre o órgão do corpo ver vesícula biliar

Uma vesícula, em biologia celular, é uma pequena estrutura dentro de uma célula, que consiste num fluido incluso por uma bicapa lipídica. Pode ser entendido como uma bolha de líquido dentro de outro líquido, ou seja, a união supramolecular constituída por diferentes moléculas. Mais tecnicamente, uma vesícula pode ser definido como um pequeno saco membranoso que pode armazenar ou transportar substâncias. As vesículas podem formar-se naturalmente devido às propriedades da sua membrana lipídica (como é o caso da micelas), ou podem ser preparadas artificialmente (como no caso dos lipossomas). A maioria das vesículas têm funções especializadas dependendo do material que contêm.
Dado que as vesículas têm um aspecto muito parecido, é muito difícil descrever a diferença entre os diferentes tipos.
A vesícula está separada do citosol pelo menos por uma bicamada de fosfolípidos. Quando possuem apenas uma destas bicamadas designam-se de unilamelares, e se não, multilamelares.
As vesículas transportam ou digerem produtos celulares ou resíduos. A membrana que rodeia a vesícula é similar à da membrana plasmática, pelo que as vesículas podem facilmente fundir-se com a membrana plasmática e libertar o seu conteúdo para fora da célula. As vesículas podem também fundir-se com outros organelos da célula.
Como o interior da vesícula está separado do citosol pela sua membrana, o interior pode ser diferente do ambiente citosólico. Graças a isso, as vesículas são a ferramenta básica celular para a organização das substâncias celulares. As vesículas estão envolvidas no metabolismo, transporte, controlo de flutuação, e armazenamento de enzimas. Podem também desempenhar o papel de compartimentos para a formação de reações químicas.
Tipos de vesículas
Vacúolos
Os vacúolos são vesículas que contém principalmente água.
- As células vegetais distinguem-se por ter um grande vacúolo central que ocupa a maior parte do volume celular, e que a célula da planta usa para o controlo osmótico e o armazenamento de nutrientes, mas também pode armazenar outros produtos e possui enzimas.
- Os vacúolos alimentares intervêm na fagocitose e em outras formas de endocitose. As vesículas de endocitose recebem geralmente o nome de endossomas.
- Os vacúolos contracteis são encontrados em determinados protistas, especialmente entre os cilióforos. Esses vacúolos recolhem água do citoplasma e expelem-na para fora da célula por um poro especial, evitando assim rebentar pela pressão osmótica.
Lisossomas
- Os lisossomas intervêm na digestão celular. O alimento que é fagocitado do exterior e fica incluído num vacúolo alimentício por endocitose é digerido pelas enzimas lisossómicas quando o vacúolo se funde com o lisossoma, dando origem a um vacúolo digestivo. Estão também envolvidos na destruição de microorganismos fagocitados pelos glóbulos brancos do sangue, e no processamento de substâncias dentro da célula, em associação com os endossoma s.
- Os lisossomas também são usados para destruir organelos celulares defeituosos ou danificados, processo este denominado endofagocitose.
Vesículas de transporte
- As vesículas de transporte podem transportar moléculas entre duas zonas do interior da célula, por exemplo, deslocam proteínas desde o retículo endoplasmático rugoso ao aparelho de Golgi.
- As proteínas de membrana e as proteínas para a secreção sintetizam-se nos ribossomas do retículo endoplasmático rugoso. A maioria dessas proteínas amadurecem no aparelho de Golgi antes de alcançarem o seu destino final, que pode ser um lisossoma, um peroxissoma ou o exterior da célula. Estas proteínas viajam no interior da célula dentro de vesículas de transporte.
Vesículas secretoras
As vesículas secretoras contêm substâncias que devem ser excretadas para fora da célula. As células têm necessidade de excretar substâncias por vários motivos, nomeadamente a eliminação de resíduos ou a secreção de substâncias produzidas pela célula que desempenham uma função no organismo (hormónios, neurotransmissores, etc.), como acontece em células especializadas que armazenam esses produtos em vesículas secretoras e depois libertam-nos quando necessário. Alguns exemplos de vesículas secretoras de células especializadas são:
Tipos de vesículas secretoras
- As vesículas sinápticas localizam-se no terminal pré-sináptico de neurónios e armazenam neurotransmissores. Quando um sinal desce pelo axónio, as vesículas sinápticas fundem-se com a membrana do neurónio, libertando os neurotransmissores para que possam se ligar aos receptores de membrana do neurónio seguinte e assim se transmita o impulso nervoso.
- Nos animais os tecidos endócrinos liberam hormónios na circulação sanguínea. Estas hormonas são armazenadas primeiro a vesículas secretoras. Um bom exemplo disto é o tecido endócrino das ilhotas de Langerhans no pâncreas. Este tecido contém vários tipos celulares, os quais se definem pelas hormonas que produzem.
- As vesículas secretoras contêm enzimas que são usadas na fabricação da parede celular de plantas, protistas, fungos, bactérias e arqueas ou da matriz extracelular das células animais.
Outros tipos de vesículas
- As vesículas de gás de arqueas, bactérias e muitos microorganismos planctónicos, parece que servem para controlar a migração vertical na coluna de água, regulando o teor de gás da vesícula e, portanto, a flutuabilidade, ou permitem à célula situar-se de modo a uma captação máxima da luz.
- As vesículas da matriz extracelular que intervêm na mineralização localizam-se no espaço extracelular ou matriz do tecido cartilaginoso e ósseo. Foram descobertas independentemente em 1967 em observações com microscópio eletrónico por H. Clarke Anderson e Ermanno Bonucci. Estas vesículas exteriores derivam das células e são especializadas em iniciar a biomineralização da matriz de diversos tecidos, como de osso, cartilagem e dentina. Durante o processo normal de calcificação, um aumento do fluxo de íões cálcio e fosfato nas células é acompanhado pela apoptose da célula (autodestruição geneticamente determinada) e pela formação de vesículas da matriz. A entrada de cálcio também leva à formação de complexos de fosfatidilserina-cálcio-fosfato na membrana plasmática, em parte, mediada pelas proteínas anexinas. As vesículas da matriz evaginam-se da membrana plasmática em locais de interacção com a matriz extracelular, e transportam para a matriz extracelular cálcio, fosfato, lípidos e proteínas anexinas, que atuam para nuclear a formação de cristais do mineral. Estes processos são coordenados de maneira muito precisa para conseguir, no momento e lugar adequados, a mineralização da matriz do tecido.
- Os endossomas conhecidos como corpos multivesículares ou MVB, são vesículas revestidas por membranas contendo no seu interior outras pequenas vesículas, chamadas vesículas luminais, que se originaram por invaginação da membrana perimetral do endossoma. Se a suas vesículas internas se libertam no exterior da célula podem dar origem a exossomas.
Formação e transporte de vesículas
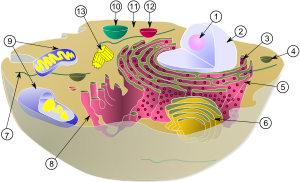
Algumas vesículas formam-se quando parte da membrana do retículo endoplasmático ou do aparelho de Golgi se evagina do organelo. Outras formam-se quando algum elemento de fora da célula é introduzido no seu interior por invaginação da membrana plasmática ou por fagocitose com emissão de pseudópodes.
Captura das moléculas a transportar
A formação das vesículas requer a fabricação de numerosos revestimentos para as estas, que as rodeiam e se unem às proteínas que transportam no seu interior. Os revestimentos também atraiem diversas proteínas receptoras, chamadas receptoras da carga, as quais ficam encarregues de capturar as moléculas da carga para transportar, e que se unirão a elas.
Revestimento de vesículas
O revestimento das vesículas serve para conformar a superfície da membrana doadora, e para seleccionar as proteínas específicas que formarão a carga para transportar na vesícula. A selecção das proteínas da carga é feita unindo-as a determinadas marcas de classificação que possuem essas proteínas. Os complexos ou clusters proteicos do revestimento da vesícula selecionam proteínas de membrana de carga nas vesículas que se evaginam.
Existem três tipos de revestimentos vesiculares, chamados coatómeros: clatrina, COPI e COPII. Os revestimentos de clatrina são comuns das vesículas que fazem o transporte entre o aparelho de Golgi e a membrana plasmática, ou entre o Golgi e os endossomas, ou entre a membrana plasmática e endossomas. As vesículas revestidas COPI são as responsáveis pelo transporte retrógrado desde o Golgi ao retículo endoplasmático. Por fim, as vesículas revestidas COPII encarregam-se do transporte anterógrado desde o retículo endoplasmático ao Golgi.
Acredita-se que o revestimento de clatrina se reconstrua em resposta à proteína reguladora G. O revestimento de coatómeros reconstrói-se e desconstrói-se devido à proteína ARF ou factor de ADP-ribosilação.
União das vesículas a membranas
Diversos marcadores de superfície das vesículas chamados SNAREs identificam qual deve ser a carga a transportar na vesícula, e complementarmente outros SNAREs da membrana do organelo de destino funcionam facilitando a fusão da vesícula transportadora com a membrana alvo. Os SNAREs vesiculares são por vezes chamados cone v-SNAREs (v de vesícula) e os da membrana alvo como t-SNARES (t de target , alvo).
Porém, não raras vezes, os SNAREs associados a vesículas ou a membranas alvo são alternadamente classificados como SNAREs Qa, Qb, Qc ou R, porque apresentam uma maior diversidade que não se poderia classificar simplesmente nos dois grupos V- ou t-SNAREs. Até agora, foram identificados em diferentes tecidos e compartimentos sub-celulares um conjunto de complexos SNARE diferentes com até 36 isoformas em humanos.
Pensa-se que as proteínas reguladoras Rab supervisionam a união dos SNAREs. As proteínas Rab são proteínas reguladoras ligadas ao GTP, e controlam a união dos SNAREs complementares durante um longo período de tempo, suficiente para que a proteína Rab hidrolize o GTP ligado e fixe a vesícula na membrana alvo.
Fusão de vesículas
Para que ocorra a fusão das vesículas com as membranas alvo é necessário que ambas se aproximem até uma distância de 1,5 nm. Para que isso ocorra, a água deve ser deslocada da superfície da membrana vesicular. Isto é energeticamente desfavorável, e diversos estudos sugerem que o processo requer ATP, GTP e acetil-CoA.
Regulação dos receptores vesiculares
As proteínas de membrana que servem como receptores são por vezes marcadas com ubiquitina para a sua eliminação e diminuição da sua quantidade. Quando chegam a um endossoma pela via anteriormente descrita, começam a formar vesículas no endossoma, que levam com elas as proteínas de membrana destinadas à degradação. Quando o endossoma matura convertendo-se em lisossoma ou quando se liga um lisossoma, as vesículas que estão no seu interior são completamente degradadas. Sem este mecanismo só a parte extracelular das proteínas de membrana chegaria ao lume do lisossoma, e só esta parte seria degradada.
Devido a estas vesículas internas que possuem endossomas, estes são denominados corpos multivesiculares. O seu meio de formação não é totalmente compreendido, e, ao contrário das outras vesículas descritas anteriormente, a superfície exterior destas vesículas não está em contacto com o citosol, uma vez que estão dentro do endossoma.
Preparação de vesículas
Têm-se realizado muitos estudos bioquímicos sobre as vesículas fosfolipídica. Para tal pode-se preparar por sonicação uma suspensão homogénea de vesículas fosfolipídica, ou injectar uma solução de fosfolípidos numa solução-tampão aquosa de membranas. Deste modo, podem preparar-se soluções aquosas de vesículas de diferentes tamanhos e com diferentes composições fosfolipídicas.
Ver também
- Retículo endoplasmático
- Aparelho de Golgi
- Micela
- Endocitose
- Vesícula sináptica
- Sistema endomembranoso
- Exossoma
- Exómero
Bibliografia
- Bruce Alberts, et al. (1994); Molecular Biology of the Cell; 3.ª Edição
Ligações externas
| Sistema endomembranoso |
 |
|
|---|---|---|
| Citoesqueleto | ||
| Endossimbiontes | ||
| Outros | ||
| Externos | ||