

As hormonas (português europeu) ou hormônios (português brasileiro) vegetais (também conhecidos como fito-hormonas (português europeu) ou fito-hormônios (português brasileiro)) são moléculas de sinalização produzidas dentro das plantas, que ocorrem em concentrações extremamente baixas. Os hormônios vegetais controlam todos os aspectos do crescimento e desenvolvimento das plantas, desde a embriogênese, a regulação do tamanho do órgão, a defesa dos agentes patogênicos, tolerância ao estresse e até o desenvolvimento reprodutivo. Ao contrário dos animais (nos quais a produção hormonal é restrita a glândulas especializadas), cada célula vegetal é capaz de produzir hormônios.Went e Thimann cunharam o termo "fito-hormônio" e o usaram no título de seu livro de 1937.
Os fito-hormônios ocorrem em todo o reino vegetal, e até mesmo nas algas, onde desempenham funções semelhantes às observadas nas plantas vasculares. Alguns fito-hormônios também ocorrem em microrganismos, como fungos e bactérias unicelulares, no entanto, nesses casos, eles não desempenham um papel hormonal e podem ser melhor considerados metabólitos secundários.
Características

A palavra hormônio é derivada do grego hormon, ou seja, excitar, colocar em movimento. Os hormônios vegetais afetam a expressão gênica e os níveis de transcrição, divisão celular e crescimento. Eles são produzidos naturalmente nas plantas, embora produtos químicos muito semelhantes sejam produzidos por fungos e bactérias que também podem afetar o crescimento das plantas. Um grande número de compostos químicos relacionados é sintetizado por seres humanos. Eles são usados para regular o crescimento de plantas cultivadas, ervas daninhas e plantas cultivadas in vitro e células vegetais; esses compostos sintéticos são chamados de reguladores de crescimento vegetal ou RCV. No início do estudo de hormônios vegetais, "fito-hormônio" era o termo comumente usado, mas seu uso é menos amplamente aplicado agora.
Hormônios vegetais não são nutrientes, mas produtos químicos que em pequenas quantidades promovem e influenciam o crescimento, desenvolvimento e diferenciação de células e tecidos. A biossíntese de hormônios vegetais nos tecidos vegetais é frequentemente difusa e nem sempre localizada. As plantas carecem de glândulas para produzir e armazenar hormônios, porque, diferentemente dos animais — que têm dois sistemas circulatórios (linfáticos e cardiovasculares) alimentados por um coração que move fluidos pelo corpo — as plantas usam meios mais passivos para mover substâncias químicas pelo corpo. As plantas utilizam produtos químicos simples como hormônios, que se movem mais facilmente através de seus tecidos. Eles são frequentemente produzidos e usados localmente dentro do corpo da planta. As células vegetais produzem hormônios que afetam até diferentes regiões da célula produtora do hormônio.
Os hormônios são transportados dentro da planta, utilizando quatro tipos de movimentos. Para o movimento localizado, utiliza-se o fluxo citoplasmático dentro das células e a lenta difusão de íons e moléculas entre as células. Os tecidos vasculares são usados para mover hormônios de uma parte da planta para outra; incluem tubos crivados ou floema que movem açúcares das folhas para as raízes e flores e xilema que move água e solutos minerais das raízes para a folhagem.
Nem todas as células vegetais respondem aos hormônios, mas essas células são programadas para responder em pontos específicos do ciclo de crescimento. Os maiores efeitos ocorrem em estágios específicos durante a vida da célula, com efeitos diminuídos antes ou depois desse período. As plantas precisam de hormônios em períodos muito específicos durante o seu crescimento e em locais específicos. Elas também precisam cortar os efeitos que os hormônios têm quando não são mais necessários. A produção de hormônios ocorre com muita frequência em locais de crescimento ativo nos meristemas, antes que as células se diferenciem totalmente. Após a produção, às vezes são movidos para outras partes da planta, onde causam um efeito imediato; ou eles podem ser armazenados nas células para serem liberados posteriormente. As plantas usam caminhos diferentes para regular as quantidades hormonais internas e moderar seus efeitos; eles podem regular a quantidade de produtos químicos usados para biossintetizar hormônios. Elas podem armazená-los nas células, inativá-los ou canibalizar hormônios já formados, conjugando-os com carboidratos, aminoácidos ou peptídeos. As plantas também podem quebrar os hormônios quimicamente, destruindo-os efetivamente. Hormônios vegetais frequentemente regulam as concentrações de outros hormônios vegetais. As plantas também movem hormônios ao redor da planta, diluindo suas concentrações.
A concentração de hormonas necessárias para respostas das plantas é muito baixa (10~6 a 10-5moles/L). Devido a essas baixas concentrações, tem sido muito difícil estudar hormônios vegetais, e somente desde o final da década de 1970 os cientistas foram capazes de começar a reunir seus efeitos e relações com a fisiologia das plantas. Grande parte do trabalho inicial sobre hormônios vegetais envolveu o estudo de plantas geneticamente deficientes em um ou envolveu o uso de plantas cultivadas em tecidos cultivadas in vitro, que foram submetidas a diferentes proporções de hormônios e o crescimento resultante foi comparado. As primeiras observações e estudos científicos datam da década de 1880; a determinação e observação dos hormônios vegetais e sua identificação foram disseminadas nos setenta anos seguintes.
Classes
Hormônios diferentes podem ser classificados em diferentes classes, dependendo de suas estruturas químicas. Dentro de cada classe de hormônio, as estruturas exatas variam, mas têm efeitos fisiológicos semelhantes. A pesquisa inicial sobre hormônios vegetais identificou cinco classes principais: ácido abscísico, auxina, citocininas, etileno e giberelinas. Esta lista foi posteriormente expandida e os brassinoesteroides, jasmonatos, ácido salicílico e estrigolactonas são agora considerados os principais hormônios vegetais. Além disso, também existem vários outros compostos que desempenham uma função semelhante aos principais hormônios, mas seu status como hormônios ósseos ainda é discutido.
Ácido abscísico
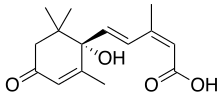
O ácido abscísico (também chamado ABA) é um dos mais importantes inibidores de crescimento das plantas. Foi descoberto e pesquisado sob dois nomes diferentes antes que suas propriedades químicas fossem completamente conhecidas, foi chamado dormina e abscicina II. Uma vez que foi determinado que os dois compostos são o mesmo, foi denominado ácido abscísico. O nome "ácido abscísico" foi dado porque foi encontrado em altas concentrações em folhas recém-abscissadas ou caídas recentemente.
Essa classe de RCV é composta por um composto químico normalmente produzido nas folhas das plantas, originário dos cloroplastos, principalmente quando as plantas estão sob estresse. Em geral, atua como um composto químico inibitório que afeta o crescimento e a dormência de sementes e brotos. Medeia mudanças no meristema apical, causando dormência dos brotos e a alteração do último conjunto de folhas em coberturas protetoras dos brotos. Uma vez que foi encontrado em folhas recém-abscissadas, acreditava-se que ele desempenha um papel nos processos de queda natural das folhas, mas pesquisas posteriores refutaram isso. Em espécies de plantas de partes temperadas do mundo, ela desempenha um papel na dormência de folhas e sementes ao inibir o crescimento, mas, como é dissipada a partir de sementes ou brotos, o crescimento começa. Em outras plantas, à medida que os níveis de ABA diminuem, o crescimento começa quando os níveis de giberelina aumentam. Sem o ABA, brotos e sementes começariam a crescer durante os períodos quentes do inverno e seriam mortos quando congelassem novamente. Como o ABA se dissipa lentamente dos tecidos e seus efeitos levam tempo para serem compensados por outros hormônios vegetais, há um atraso nas vias fisiológicas que fornecem alguma proteção contra o crescimento prematuro. Ele se acumula nas sementes durante a maturação dos frutos, impedindo a germinação das sementes dentro da fruta ou antes do inverno. Os efeitos do ácido abscísico são degradados nos tecidos das plantas durante temperaturas frias ou por sua remoção por lavagem com água dos tecidos, liberando as sementes e os brotos da dormência.
Em plantas sob estresse hídrico, o ABA desempenha um papel importante no fechamento dos estômatos. Logo depois que as plantas sofrem estresse hídrico e as raízes são deficientes em água, um sinal se move para as folhas, causando a formação de precursores ABA por lá, que depois se deslocam para as raízes. As raízes liberam o ABA, que é translocado para a folhagem através do sistema vascular e modula a captação de potássio e sódio nas células de guarda, que perdem a turgidez, fechando os estômatos. O ABA existe em todas as partes da planta e sua concentração em qualquer tecido parece mediar seus efeitos e funcionar como um hormônio; sua degradação, ou mais apropriadamente catabolismo, dentro da planta afeta reações metabólicas e crescimento celular e produção de outros hormônios. As plantas começam a vida como uma semente com altos níveis de ABA. Pouco antes de a semente germinar, os níveis de ABA diminuem; durante a germinação e crescimento inicial das mudas, os níveis de ABA diminuem ainda mais. À medida que as plantas começam a produzir brotos com folhas totalmente funcionais, os níveis de ABA começam a aumentar, diminuindo o crescimento celular em áreas mais "maduras" da planta. O estresse da água ou da predação afeta as taxas de produção e catabolismo de ABA, mediando outra cascata de efeitos que desencadeiam respostas específicas das células-alvo. Os cientistas ainda estão reunindo as complexas interações e efeitos deste e de outros fito-hormônios.
Auxinas

Auxinas são compostos que influenciam positivamente o aumento celular, formação de brotos e iniciação radicular. Elas também promovem a produção de outros hormônios e, em conjunto com citocininas, controlam o crescimento de caules, raízes e frutos e convertem os caules em flores. As auxinas foram a primeira classe de reguladores de crescimento descoberta. Elas afetam o alongamento celular alterando a plasticidade da parede celular. Elas estimulam o câmbio, um subtipo de células meristemáticas, a se dividir e, em hastes, fazem com que o xilema secundário se diferencie. As auxinas agem para inibir o crescimento dos brotos mais abaixo das hastes (dominância apical) e também para promover o desenvolvimento e o crescimento radicular lateral e adventício. A abscisão foliar é iniciada pelo ponto de crescimento de uma planta que deixa de produzir auxinas. As auxinas nas sementes regulam a síntese proteica específica, medida que se desenvolve dentro da flor após a polinização, fazendo com que a flor desenvolva um fruto para conter as sementes em desenvolvimento. As auxinas são tóxicas para as plantas em grandes concentrações; elas são mais tóxicas para as dicotiledôneas e menos para as monocotiledôneas. Devido a essa propriedade, foram desenvolvidos e utilizados herbicidas de auxina sintética, incluindo 2,4-D (2,4-diclorofenoxiacético) e 2,4,5-T, para o controle de ervas daninhas. As auxinas, especialmente o ácido 1-naftalenacético (NAA) e o ácido indol-3-butírico (IBA), também são comumente aplicadas para estimular o crescimento radicular quando se tomam estaquias de plantas. A auxina mais comum encontrada nas plantas é o ácido indolacético ou IAA.
Brassinosteróides

Os brassinosteroides são uma classe de poli-hidroxiesteroides, o único exemplo de hormônios baseados em esteroides nas plantas. Os brassinosteroides controlam o alongamento e a divisão celular, o gravitropismo, a resistência ao estresse e a diferenciação do xilema. Eles inibem o crescimento radicular e a abscisão foliar. O brassinolida foi o primeiro brassinosteroide identificado e foi isolado a partir de extratos de pólen de colza (Brassica napus) em 1979.
Citocininas
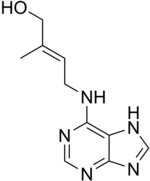
As citocininas são um grupo de produtos químicos que influenciam a divisão celular e a formação de brotações. Eles eram chamados cininas no passado, quando as primeiras citocininas foram isoladas de células de levedura. Elas também ajudam a retardar a senescência dos tecidos, são responsáveis pela mediação do transporte de auxinas por toda a planta e afetam o comprimento internodal e o crescimento das folhas. As citocininas e auxinas geralmente trabalham juntas, e as proporções desses dois grupos de hormônios vegetais afetam a maioria dos principais períodos de crescimento durante a vida útil da planta. As citocininas combatem a dominância apical induzida pelas auxinas; Em conjunto com o etileno, promove a abscisão de folhas, partes de flores e frutos.
Etileno
O etileno é um gás que se forma através da decomposição da metionina, que está presente em todas as células. O etileno tem uma solubilidade muito limitada na água e não se acumula dentro da célula, mas se difunde para fora da célula e escapa para fora da planta. Sua eficácia como hormônio vegetal depende da taxa de produção versus a taxa de fuga para a atmosfera. O etileno é produzido a uma taxa mais rápida nas células que crescem e se dividem rapidamente, especialmente no escuro. Novo crescimento e plântulas recém-germinadas produzem mais etileno do que o que pode escapar da planta, o que leva a quantidades elevadas de etileno, inibindo a expansão foliar. À medida que o novo broto é exposto à luz, reações mediadas pelo fitocromo, as células da planta produzem um sinal para a produção de etileno diminuir, permitindo a expansão das folhas. O etileno afeta o crescimento e o formato das células; quando um rebento ou raiz em crescimento atinge um obstáculo enquanto está no subsolo, a produção de etileno aumenta muito, impedindo o alongamento celular e causando o inchamento do caule. A haste mais grossa resultante é mais forte e menos propensa a dobrar sob pressão à medida que pressiona contra o objeto, impedindo seu caminho para a superfície. Se a parte aérea não atingir a superfície e o estímulo ao etileno se prolongar, isso afeta a resposta geotrópica natural do caule, que deve crescer na vertical, permitindo que ela cresça ao redor de um objeto. Estudos parecem indicar que o etileno afeta o diâmetro e a altura do caule: quando as hastes das árvores são submetidas ao vento, causando estresse lateral, ocorre uma maior produção de etileno, resultando em troncos e galhos de árvores mais espessos e mais resistentes. O etileno afeta o amadurecimento das frutas: normalmente, quando as sementes estão maduras, a produção de etileno aumenta e se acumula no fruto, resultando em um evento climatérico imediatamente antes da dispersão das sementes. A proteína nuclear Ethylene Insensitive2 (EIN2) é regulada pela produção de etileno e, por sua vez, regula outros hormônios, incluindo o ABA e os hormônios do estresse. A difusão de etileno das plantas é fortemente inibida debaixo d'água. Isso aumenta as concentrações internas do gás. Em inúmeras espécies aquáticas e semiaquáticas (por exemplo, Callitriche platycarpus, arroz e Rumex palustris), o etileno acumulado estimula fortemente o alongamento ascendente. Essa resposta é um mecanismo importante para a fuga adaptativa da submersão que evita a asfixia retornando a parte aérea e deixando o contato com o ar, permitindo a liberação de etileno retido. Pelo menos uma espécie (Potamogeton pectinatus) foi considerada incapaz de produzir etileno, mantendo uma morfologia convencional. Isso sugere que o etileno é um verdadeiro regulador, e não um requisito para a construção do plano básico do corpo de uma planta.
Giberelinas

As giberelinas (GAs) incluem uma grande variedade de produtos químicos produzidos naturalmente nas plantas e pelos fungos. Elas foram descobertas pela primeira vez quando pesquisadores japoneses, incluindo Eiichi Kurosawa, notaram uma substância química produzida por um fungo chamado Gibberella fujikuroi que produzia crescimento anormal nas plantas de arroz. Mais tarde, foi descoberto que as GAs também são produzidas pelas próprias plantas e controlam vários aspectos do desenvolvimento ao longo do ciclo de vida. A síntese da GA é fortemente regulada em sementes na germinação e sua presença é necessária para que a germinação ocorra. Em mudas e adultos, as GAs promovem fortemente o alongamento celular. As GAs também promovem a transição entre crescimento vegetativo e reprodutivo e também são necessárias para a função do pólen durante a fertilização.
Jasmonatos

Os jasmonatos (JAs) são hormônios lipídicos que foram originalmente isolados do óleo de jasmim. Os JAs são especialmente importantes na resposta das plantas ao ataque de herbívoros e patógenos necrotróficos. O JA mais ativo nas plantas é o ácido jasmônico. O ácido jasmônico pode ser metabolizado ainda mais em metil-JA, que é um composto orgânico volátil. Essa propriedade incomum significa que o metil-JA pode atuar como um sinal aéreo para comunicar o ataque de herbívoros a outras folhas distantes dentro de uma planta e até como um sinal para as plantas vizinhas. Além de seu papel na defesa, acredita-se que os JAs também desempenhem papéis na germinação de sementes, no armazenamento de proteínas nas sementes e no crescimento radicular.
Ácido salicílico

O ácido salicílico (AS) é um hormônio com uma estrutura relacionada ao fenol. Foi originalmente isolado de um extrato de casca de salgueiro-branco (Salix alba) e é de grande interesse para a medicina humana, pois é o precursor do analgésico aspirina. Nas plantas, o AS desempenha um papel crítico na defesa contra patógenos biotróficos. De maneira semelhante aos JAs, o AS também pode se tornar metilado. Como o metil-JA, o metil-AS é volátil e pode atuar como um sinal de longa distância para as plantas vizinhas para alertar sobre o ataque de patógenos. Além de seu papel na defesa, o AS também está envolvido na resposta das plantas ao estresse abiótico (particularmente seca, temperatura, metais pesados e estresse osmótico).
Estrigolactonas

As estrigolactonas foram originalmente descobertas através de estudos sobre a germinação da erva parasita Striga lutea. Verificou-se que a germinação das espécies Striga foi estimulada pela presença de um composto exsudado pelas raízes da planta hospedeira. Mais tarde, foi demonstrado que as estrigolactonas que são exsudadas no solo promovem o crescimento de fungos micorrízicos arbusculares simbióticos. Mais recentemente, outro papel das estrigolactonas foi identificado na inibição da ramificação da parte aérea. Essa descoberta do papel das estrigolactonas na ramificação de brotações levou a um aumento dramático no interesse desses hormônios, e desde então foi demonstrado que as estrigolactonas desempenham papéis importantes na senescência foliar, resposta à falta de fosfato, tolerância ao sal e sinalização luminosa.
Outros hormônios conhecidos
Outros reguladores de crescimento de plantas identificados incluem:
- Hormônios peptídicos vegetais — engloba todos os pequenos peptídeos secretados envolvidos na sinalização célula a célula. Esses pequenos hormônios peptídicos desempenham papéis cruciais no crescimento e desenvolvimento das plantas, incluindo mecanismos de defesa, controle da divisão e expansão celular e auto-incompatibilidade do pólen. Sabe-se que o pequeno peptídeo CLE25 atua como um sinal de longa distância para comunicar o estresse hídrico detectado nas raízes aos estômatos das folhas.
- Poliaminas — são moléculas fortemente básicas com baixo peso molecular que foram encontradas em todos os organismos estudados até o momento. Elas são essenciais para o crescimento e desenvolvimento das plantas e afetam o processo de mitose e meiose. Nas plantas, as poliaminas têm sido associadas ao controle da senescência e à morte celular programada.
- Óxido nítrico (ON) — serve como sinal nas respostas hormonais e de defesa (por exemplo, fechamento estomático, desenvolvimento radicular, germinação, fixação de nitrogênio, morte celular, resposta ao estresse). O ON pode ser produzido por um ON sintase ainda indefinida, um tipo especial de nitrito redutase, nitrato redutase, citocromo c oxidase mitocondrial ou processos não enzimáticos e regular as funções das organelas celulares das plantas (por exemplo, síntese de ATP em cloroplastos e mitocôndrias).
- Carriquinas — não são hormônios vegetais, pois não são produzidas pelas próprias plantas, mas são encontradas na fumaça da queima de material vegetal. As carriquinas podem promover a germinação de sementes em muitas espécies. A descoberta de que as plantas que não possuem o receptor de carriquina apresentam vários fenótipos de desenvolvimento (maior acúmulo de biomassa e maior sensibilidade à seca) levou alguns a especular sobre a existência de um hormônio endógeno semelhante à carriquina ainda não identificado nas plantas. A via de sinalização de carriquina celular compartilha muitos componentes com a via de sinalização de estrigolactona.
- Triacontanol — um álcool graxo que atua como estimulante do crescimento, iniciando especialmente novas quebras basais na família das rosas. Pode ser encontrado na alfafa (lucerna), cera de abelha e algumas cutículas de folhas de cera.
Hormônios e propagação de plantas
Hormônios vegetais sintéticos ou RCV são usados em várias técnicas diferentes que envolvem a propagação de plantas a partir de estaquias, enxertia, micropropagação e cultura de tecidos.
A propagação de plantas por estacas de folhas, caules ou raízes totalmente desenvolvidas é realizada por jardineiros que utilizam a auxina como composto de enraizamento aplicado à superfície de corte; as auxinas são levadas para a planta e promovem o início da raiz. No enxerto, a auxina promove a formação de tecido caloso, que une as superfícies do enxerto. Na micropropagação, diferentes RCV são usados para promover a multiplicação e o enraizamento de novas plântulas. No cultivo de células vegetais, os PGRs são usados para produzir crescimento, multiplicação e enraizamento de calos.
Dormência de sementes
Os hormônios vegetais afetam a germinação e a dormência das sementes, agindo em diferentes partes da semente.
A dormência do embrião é caracterizada por uma alta relação ABA:GA, enquanto a semente apresenta alta sensibilidade ao ácido abscísico e baixa sensibilidade à GA. Para liberar a semente desse tipo de dormência e iniciar a germinação, uma alteração na biossíntese e degradação hormonal em direção a uma baixa relação ABA/GA, juntamente com uma diminuição na sensibilidade à ABA e um aumento na sensibilidade à GA, devem ocorrer.
A ABA controla a dormência do embrião e a germinação do embrião GA. A dormência do revestimento de sementes envolve a restrição mecânica do revestimento de sementes. Isso, junto com um baixo potencial de crescimento embrionário, produz efetivamente a dormência das sementes. A GA libera essa dormência aumentando o potencial de crescimento do embrião e/ou enfraquecendo o revestimento das sementes, para que o radical da muda possa romper o revestimento. Diferentes tipos de casacos de sementes podem ser constituídos por células vivas ou mortas, e ambos os tipos podem ser influenciados por hormônios; os compostos de células vivas são acionados após a formação das sementes, enquanto os casacos de sementes compostos de células mortas podem ser influenciados por hormônios durante a formação do revestimento das sementes. O ABA afeta as características de crescimento de testa ou revestimento de sementes, incluindo espessura, e afeta o potencial de crescimento de embriões mediado pela GA. Essas condições e efeitos ocorrem durante a formação da semente, geralmente em resposta às condições ambientais. Os hormônios também medeiam a dormência do endosperma: o endosperma na maioria das sementes é composto de tecido vivo que pode responder ativamente aos hormônios gerados pelo embrião. O endosperma geralmente atua como uma barreira à germinação das sementes, desempenhando um papel na dormência do revestimento das sementes ou no processo de germinação. As células vivas respondem e também afetam a relação ABA:GA e medeiam a sensibilidade celular. A GA aumenta assim o potencial de crescimento do embrião e pode promover o enfraquecimento do endosperma. A GA também afeta os processos independentes e inibidores de ABA no endosperma.
Aplicações em humanos
Ácido salicílico
A casca-de-salgueiro é usada há séculos como analgésico. O ingrediente ativo da casca-de-salgueiro que fornece esses efeitos é o hormônio ácido salicílico (AS). Em 1899, a empresa farmacêutica Bayer começou a comercializar um derivado do AS como o medicamento aspirina. Além de seu uso como analgésico, o AS também é usado em tratamentos tópicos de várias doenças da pele, incluindo acne, verrugas e psoríase. Foi encontrado outro derivado de AS, salicilato de sódio, para suprimir a proliferação de células cancerígenas humanas de leucemia linfoide, próstata, mama e melanoma.
Ácido jasmônico
O ácido jasmônico (JA) pode induzir a morte em células de leucemia linfoblástica. Foi demonstrado que o jasmonato de metila (um derivado do JA, também encontrado em plantas) inibe a proliferação em várias linhas de células cancerígenas, embora ainda exista um debate sobre seu uso como um medicamento anticâncer, devido aos seus potenciais efeitos negativos nas células saudáveis.
Notas
Ligações externas
- Tabela simples de hormônios vegetais com localização da síntese e efeitos da aplicação — este é o formato usado nas descrições nos finais dos artigos da Wikipédia sobre hormônios vegetais individuais.
- Regulação hormonal da expressão e desenvolvimento de genes — Introdução detalhada, incluindo informações genéticas.